Bone identification 101
The aim of this section is to demystify the process of identifying bones. It provides concrete methods that can be used to identify bone on Indigenous and historic sites across Australia. At the same time it explains the broad anatomical differences between marsupial and placental mammals. This section also briefly outlines the major components of bone, the effect of post-depositional processes on bone preservation, and some guidelines on cleaning and storing bone from archaeological contexts.
The major components of bone
Bone has both an organic (collagen and non-collagenous proteins) and inorganic (hydroxyapatite) component. The organic component (30%) is the first to disappear after the death of an organism. This explains why most archaeological bones have a lighter, brittle feeling in comparison with fresh bone. There are two main types of bone, cortical or lamellar bone and trabecular or cancellous bone. Cortical bone is found in the shafts (diaphyses) of long bones, as well as in many flat bones, like the pelvis, scapula and skull. Trabecular bone (sometimes called spongy bone) is found in the ends (epiphyses) of long bones, and functions as a cushion of sorts for the body’s joints. Morphological differences between the two types of bone have direct bearing on their preservation in archaeological contexts. For example, trabecular bone is very porous, and generally less dense than cortical bone. For this reason, when carnivores, like a dingo, chew a bone, they tend to destroy the ends. Similarly, the cortical bone, in the form of bone shafts, is generally left behind because it is denser and hardier than the epiphyses. These shafts are frequently splintered – a result of marrow extraction by carnivores (and humans), or an effect of weathering. Understanding this relationship may help identify some of the processes affecting the site and its assemblage, leading to more accurate final site interpretations.
First steps in the identification process
Identification of an unknown bone can seem to be a daunting task. When starting this process it is helpful to consider that form is a reflection of function. For example, an animal that walks on four legs (quadruped) uses each limb to support a heavy head and body. Therefore, arms and legs will be similarly sized, as both fore limbs and hind limbs are used to support the animal’s weight. However, in a biped (in this book, humans and macropods), the arms or fore limbs are used mainly for carrying and fine manipulation, and, while these may also be used for support, it is to a lesser extent than in dedicated quadrupeds. This translates into fore limb/arm bones that are generally smaller than the hind limb/leg bones. Similarly, bones of the cranium (head), pelvis (hips) and scapula (shoulder blade) are often referred to as flat bones; again, this reflects their function as more than strictly weight bearing. Vertebrae, carpals and tarsals are all classed as irregular bones, and have varying functions in the spine, wrist and ankle, respectively. Lastly, the relative thickness and/or robustness of all bones can be a further useful indicator of general species size and skeletal element. The general size and shape of a bone can be a first step in the identification of both elements and species, and is a good basic way to narrow down a potentially large number of elements into a manageable list.
While size can be a useful way in which to make sense of what can initially seem to be an impossibly large task, we must also ascertain whether the bones in question are from an adult or a juvenile. The study of juvenile and neonatal skeletal elements is a large and complicated subject, therefore this manual will touch on just some of the main issues surrounding their identification.
Juvenile skeletal elements are difficult to identify for a variety of reasons. First, the morphology of juvenile bones may differ from adult bones because these bones may not be fully fused. For example, in juveniles the tibia is initially comprised of three bones – proximal epiphysis, shaft, and distal epiphysis – that eventually fuse together to form one bone as it grows (Figures 0.3a and b). The majority of bones in the body undergo a similar process, and the rate of fusion for each bone is unique and varies by species. While this can be very helpful in determining an age at death for a particular individual, juvenile bones can make identification difficult because unfused portions of a bone may be missing in archaeological assemblages, leaving just the shaft or other fragmentary portions. When this occurs, juvenile shafts may be mistaken for other skeletal elements, and mistakenly compared with an adult bone.
The second issue in the identification of juvenile bones is the likelihood that they will not survive as well as adult bones due to their lower bone density. This means that they may be either fragmentary, or disappear altogether, in contexts where adult bone survives. Given the difficulties, juvenile bone identification should be made by a specialist and with the help of a skeletal reference collection that includes bones from immature individuals.
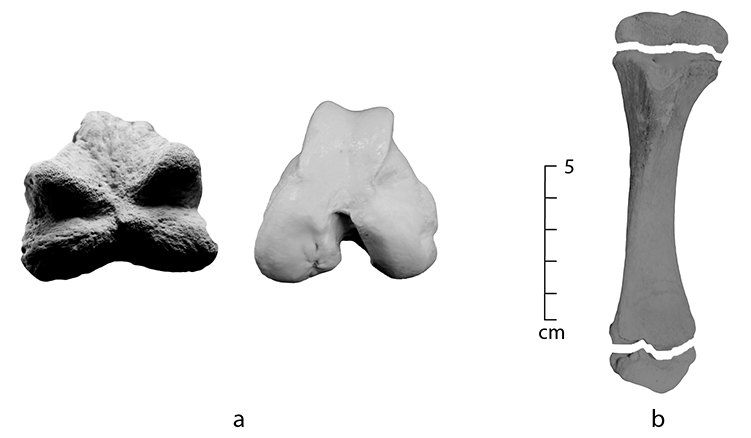
Figure 0.3: (a) A distal left pig femur with an unfused epiphysis on the left, compared to the distal end of an adult pig on the right, (b) an unfused juvenile pig tibia.
Species included
Generally, the first and arguably most important question accompanying the discovery of bone on many archaeological sites, is whether it is human or animal. This is followed by whether the bone is from an introduced or native species. Given these basic divisions, the following 16 species have been included with a view to encompassing the human–animal and the native–introduced species distinctions, as well as the historic–Indigenous site distinction.
Native species
|
Kangaroo |
Elements from an eastern grey kangaroo will permit broad identification of a bone as belonging to a large macropod. Finer distinctions between sub-species are often difficult to make on the basis of post-cranial (from the neck down) elements alone. Kangaroos occur in both historic and Indigenous contexts, although they are generally more common in the latter. |
|
Wallaby |
Red-necked wallabies may be found in both historic and Indigenous contexts. As with kangaroos, finer distinction between sub-species must rest with a specialist. |
|
Wombat |
The common wombat is similar morphologically to the southern hairy-nosed wombat. Wombats are more common in Indigenous contexts. |
|
Possum |
A brushtail possum is similar morphologically to a ringtail possum. Possums may occur in a variety of contexts. |
|
Quoll |
A tiger quoll (often referred to as a native cat) is included in this manual as an example of one of Australia’s few native carnivorous marsupials. |
|
Bandicoot |
A long-nosed bandicoot (Perameles nausuta) is included due to its ubiquity in many archaeological sites, especially Indigenous sites in north-west Australia. |
|
Emu |
Post-cranial elements of emus may be found in Indigenous contexts. Smaller wing bones are rarely recovered in archaeological contexts. |
Introduced species
|
Human |
In most cases, the most significant question asked when bone is found is whether it is human or animal. For this reason, human elements have been included for comparison. |
|
Dingo/dog |
While often viewed as a native species, the dingo was introduced to Australia during the Holocene. Morphologically, domestic dogs and dingoes are nearly indistinguishable, with the latter having slightly more robust cranial features and teeth. Distinction between the two necessitates examination by a specialist. In prehistoric contexts, dingoes may also be mistaken for thylacines (the extinct marsupial wolf) due to their morphological similarity. |
|
Sheep |
As sheep are more common than goats on most Australian historical sites, sheep bones have been depicted throughout this manual. While these elements can also be used to identify goat, the distinction between the two can be difficult. A specialist and a comparative collection should be consulted for distinction. |
|
Cow (Bos taurus), Pig (Sus scrofa) and Rabbit (Oryctolagus sp) |
Each of these species is a common food animal in Australian historic assemblages. |
|
Chicken (domestic fowl) |
In general, bird bones are relatively easy to identify due to their distinctive morphology and comparatively lighter bone weight. Chickens and domestic fowl are relatively common in urban historical assemblages. |
|
Horse/donkey |
Horses generally appear in rather low frequencies in historical assemblages. They are morphologically similar to donkeys and, due to their size, may be mistaken for cow. Cattle and horse are depicted side by side in this manual to illustrate their morphological differences. |
|
Cat |
Cats may have entered the archaeological record due to post-depositional commensal activity and are frequently encountered as intentional burials, in foundation deposits and in wells. |
The species recovered from most archaeological contexts in Australia can be quite variable, and this variability is greatly dependent on the geographic region and environment in which the assemblage is located. This manual therefore contains those native and introduced species most commonly found in both historic and prehistoric assemblages. Birds, with the exception of emus and chickens, as well as rodents and fish, have been excluded. There are several good references that cover these species, and a ‘field’ manual quickly becomes a large reference book if every species of animal were to be included.
With respect to size, the species used in this manual are grouped under the general umbrella terms ‘small’ ‘medium’ and ‘large’ to help with the identification process. These are relative terms, so a guide has been included to avoid ambiguity (Figure 0.4). Nevertheless, it is important to consider individual variability on size. For example in this framework, an adult human can range from medium to large, as can a kangaroo and emu. Do not use this framework if juvenile bones from any species are suspected.
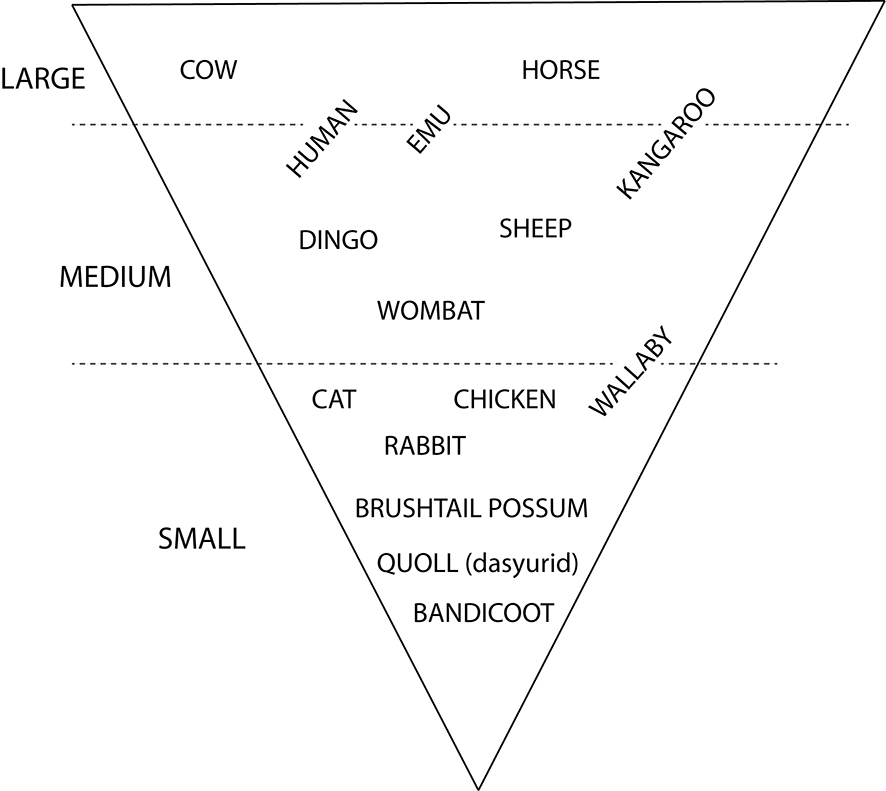
Figure 0.4: Overview of the small, medium and large animal groupings used in this manual.
Skeletal elements included
For ease of use and practicality, only selected elements are depicted in this manual. Emphasis is placed on bones that are both found in archaeological assemblages in Australia with some frequency and can be identified to species by a non-specialist, especially if incomplete.
This manual emphasises the post-cranial skeleton (neck down). However, mandibles have been included because they are commonly recovered. Teeth have also been given brief attention as part of the mandible, because they are frequently well preserved and can provide a definitive species determination, complete with age and sometimes sex. Given the specialised nature of teeth, they are a topic unto themselves, and for this reason have not been addressed in greater detail by this manual (for more information, see Hillson 2005). Crania (skulls) have also been omitted, as often they are too fragmented to be identified in the field. Mandibles often survive much better than other parts of the crania. The mandibular hinge, coupled with any in situ teeth, renders mandibles highly diagnostic elements for species identification.
The post-cranial skeleton can be sub-divided into axial (trunk), appendicular (limb) and manus/pes (hand/foot) elements. This sub-division is a useful distinction when addressing economic and subsistence-based questions, such as butchery and consumption.
The axial skeleton is comprised of vertebrae, ribs, pelvis and scapula (sometimes included in the appendicular skeleton). Due to the difficult nature of identifying ribs and vertebrae to species, this manual has omitted these elements. Their identification necessitates the use of a specialist and a good comparative collection. However, the pelvis and scapula are both elements that are highly identifiable to species, and in the case of the pelvis, sex designation may also be possible. For these reasons both the scapula and pelvis have been included.
The appendicular skeleton is comprised of the bones of the fore limb/arm (humerus, radius, ulna) and hind limb/leg (femur, tibia, fibula). With the exception of the fibula, these bones are frequently identifiable, even when fragmentary. The fibula has been excluded from this manual because although often present in archaeological assemblages, its fragmentary nature makes identification to species difficult.
The post-cranial skeleton is also comprised of manus (hand) and pes (foot) bones. As with most of the axial skeleton, these bones are more difficult to identify without the aid of a comparative collection. For this reason, the foot is treated as a distinct entity, and is depicted articulated. Several elements are highlighted in greater detail, namely the calcaneus and astragalus, because they are frequently encountered, often complete, and easily diagnostic to species.
As is evident by those bones included and omitted, this manual is not intended to provide all the answers, nor can it replace the experience of a professional zooarchaeologist or forensic osteologist. Rather it is intended as an introductory guide that will facilitate initial distinctions between human and animal, and introduced and native species. Nearly every bone can be identified at some level, especially given recent advances in molecular biology (DNA). That said, the preciseness of identification is often a result of a need to balance time, money and research questions, and this is where specialist expertise may be necessary.
Anatomical differences between marsupial and placental mammals
An important component of this manual is the depiction of both native (often marsupials) and introduced (placental) species commonly found in Australian archaeological sites. Distinguishing between these two broad groups is a lot easier with a cursory understanding of some fundamental morphological differences. The most visible difference between marsupial (metatherian) and placental (eutherian) mammals is the presence of a pouch in the former, in which immature young remain while they mature. Moreover, there are a number of morphological differences between the two that are often visible on the skeleton. The following are some general morphological variations that may be useful in distinguishing between marsupials and placentals, and thus native and introduced animals. Further specific differences are discussed in the relevant identification chapters.
Mandible
Typically the rear portion of the mandible is turned inward in marsupials (inflected jaw angle), as opposed to outward in placentals. This ‘angular process’ forms a pronounced shelf that projects medially from the posterior mandible (Figure 0.5).
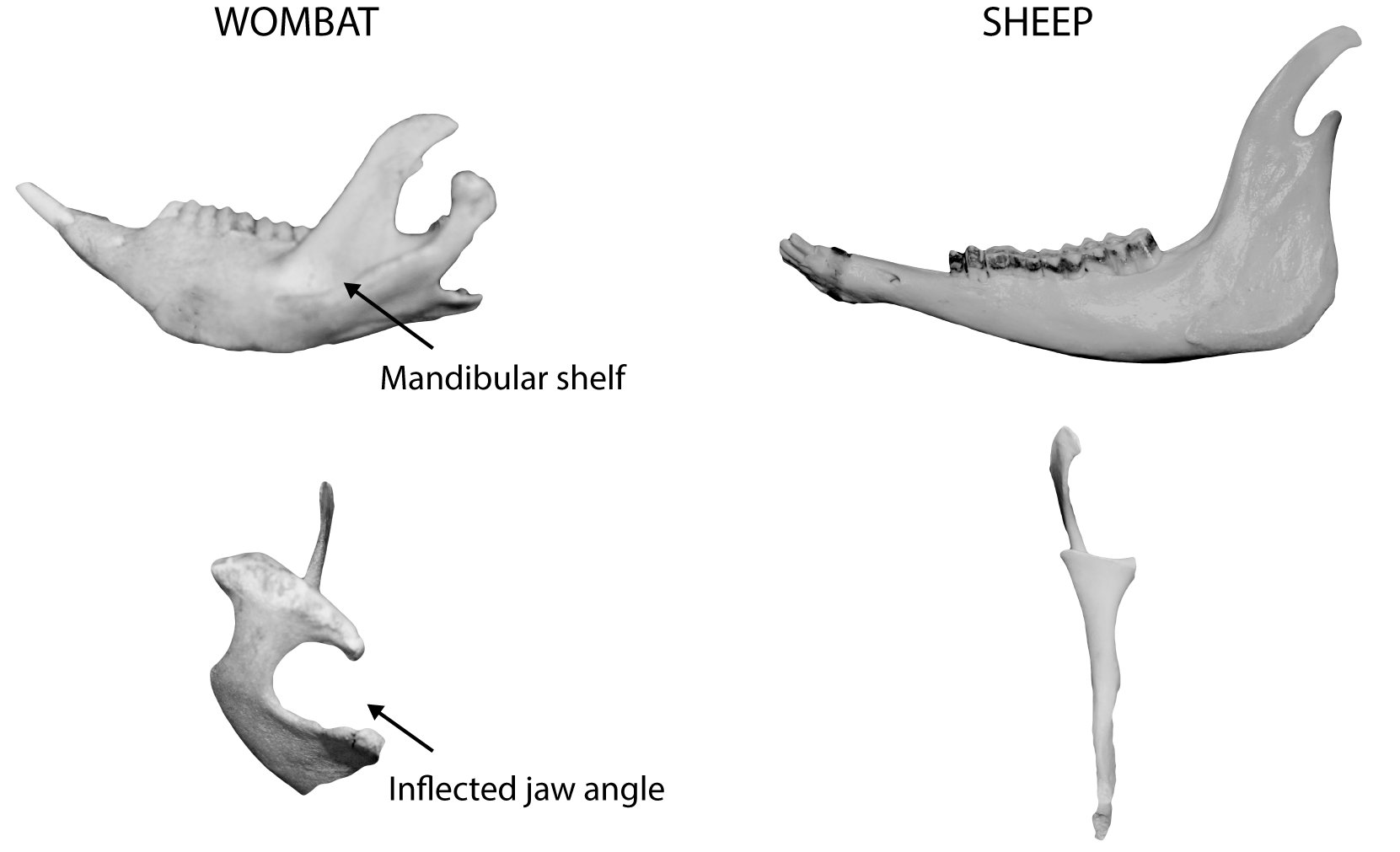
Figure 0.5: Comparison of a wombat and sheep left mandible showing the characteristic mandibular shelf and inflected jaw angle of a marsupial.
Dentition
In general, marsupials have different dental patterns than placental mammals, often having three premolars and four molars on each side in both the upper and lower jaw (P3/3; M4/4). In placental mammals, the general dental formula is four premolars and three molars on either side in both the upper and lower jaw (PM 4/4; M 3/3). The third premolar is the only deciduous tooth in marsupials, leading to what is often termed ‘molar progression’, whereby it is eventually lost later in life. The presence or absence of the third premolar may be used in age estimates.
Pelvis
Nearly all marsupials have two epipubic bones, which extend outward from the front of the pelvis (anterior pelvic symphysis) (Figure 0.6). Although some have argued that their function is unresolved, it is commonly thought they act either as support for the pouch and/or the stomach muscles. These bones generally disarticulate from the rest of the pelvis after deposition and may not be recovered.
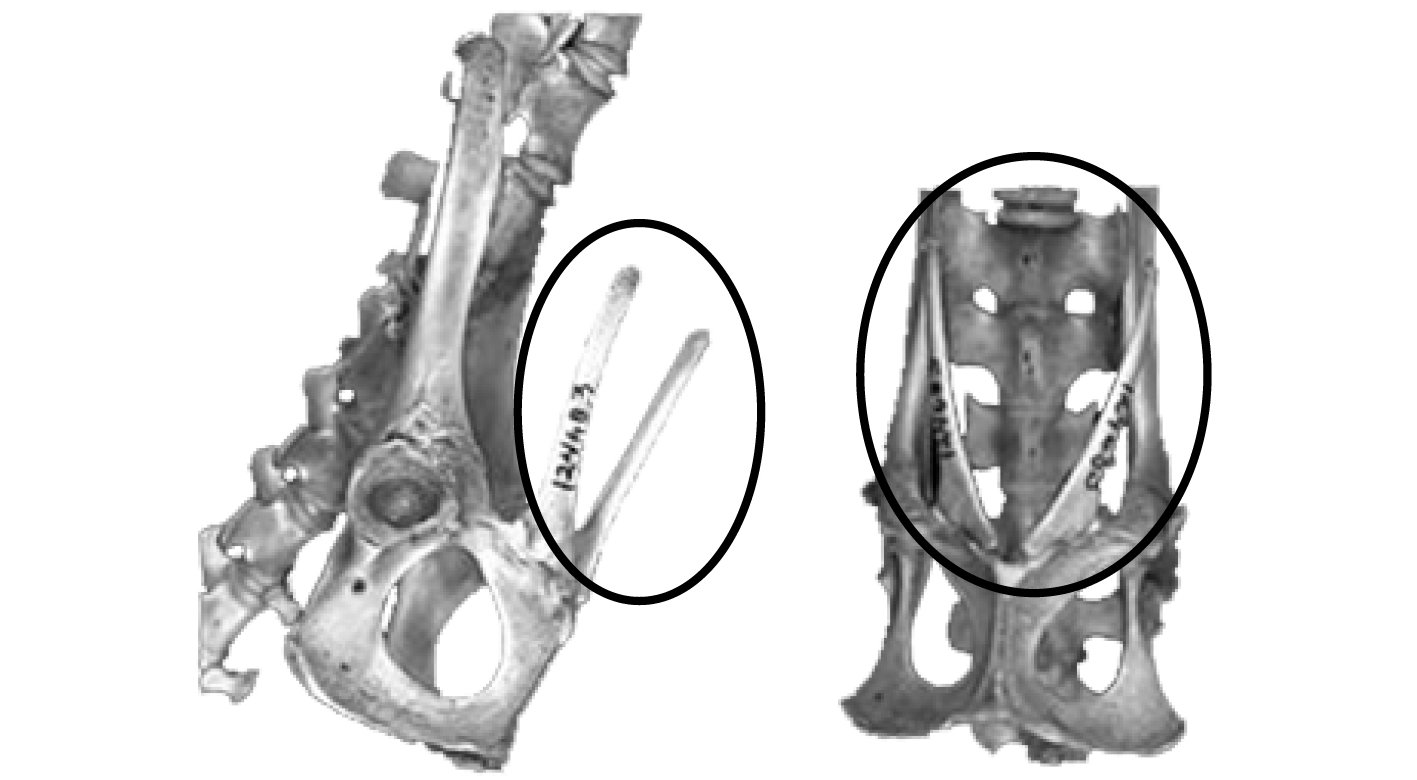
Figure 0.6: An articulated possum pelvis showing the epipubic bone (circled).
Flared humerus
The medio-distal humerus of most marsupials is characterised by marked flaring, often culminating in a sharp crest found on the lateral side, which ‘points’ up to the proximal end. This crest is balanced by an epicondylar foramen on the medial side (Figure 0.7).
Epiphyseal fusion
In marsupials, the proximal tibia and proximal humerus often fail to fully fuse.
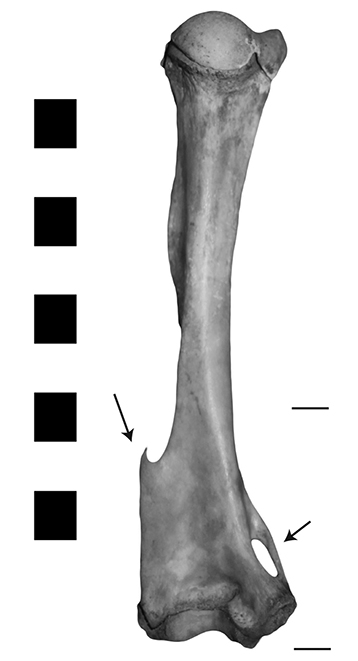
Figure 0.7: A kangaroo humerus showing the flared medio-distal portion, and the lateral crest and epicondylar foramen (arrows).
In general, consideration of the environment in which the species are found can be an important aid in the initial differentiation between marsupial and placental (or native and introduced) species, providing valuable clues about the types of animals you can expect to recover. For example, the species found in an urban domestic rubbish pit would vary greatly from those you might encounter during a survey in the Pilbara. While there are exceptions to this generalisation, any information that can help to narrow down the potential list of species is useful.
Post-depositional processes (taphonomy)
Taphonomy is the study of everything that happens to an individual after death. Understanding the range of factors that could have affected bones after they were deposited is necessary to provide a full interpretation of an archaeological site. There are many taphonomic factors that could exert an influence on bones in an assemblage, and this manual will prioritise those that are easily identifiable for a non-specialist, and most relative to Australian contexts (for a detailed discussion of taphonomy, see Lyman 1994).
Factors that can affect bone after an organism’s death can be loosely grouped into two categories: natural and anthropogenic. Modifications resulting from natural causes include scavenging by other species (Figures 0.8a and b), as well as extremes of heat and cold (Figure 0.8c), floods, drought and soil chemistry. As with any once living thing, environmental changes, such as fluctuations in temperature and moisture, are enemies of long-term preservation. Other natural factors affecting preservation include movement by wind or water, or trampling by animals. These types of movement can scatter assemblages and leave tell-tale abrasions on bone surfaces that may be visible under a microscope (and occasionally by the naked eye). Animals such as feral pigs and dingoes also scavenge carcasses, and in addition to leaving tooth marks on bones (Figures 0.8a and b), may destroy and scatter elements.

Figure 0.8: Bone modifications from natural and anthropogenic causes: (a) the distal end of a gnawed rib, (b) a bone heavily gnawed by a rat, (c) a bone showing characteristic cracking and flaking from exposure, and (d) a bone sawing through the shaft, illustrating the distinctive marking that results from a band saw.
Anthropogenic factors affecting preservation can include cut marks resulting from butchery. These may be quite subtle, or in the case of historic period abattoirs, quite obvious, such as the distinctive striations left by butcher’s band saws (Figure 0.8d). Human agency is also a key factor in skeletal element patterning in archaeological assemblages. For example, an assemblage dominated by limb bones may suggest the remains of dietary refuse (as these elements typically contain a lot of meat). If this same assemblage does not contain cranial or foot bones, we could further suggest that initial butchery took place elsewhere, with the limb bones subsequently transported for sale and consumption. Conversely, if relatively complete skeletons are recovered, on-site butchery or death may have occurred. Humans may also cook bone, and so elements can bear traces of burning and/or boiling. Industrial activities are also a possibility, and these include glue and tallow manufacturing, along with tanneries and craft workshops. Recent advances in DNA have now made it possible to potentially identify the species from which worked bone objects, such as buttons and beads, originally came.
Post-excavation cleaning and storage
Fluctuations in temperature and/or humidity, and insect activity are major issues in bone preservation following excavation. Therefore, the best way to store bone is in a dry, cool environment that is free from insects and rodents (who like to gnaw on dry bone). Bone should be cleaned after excavation with a soft, dry brush. A little water may be used if necessary. Harsh chemicals and detergents should be avoided, as well as stiff brushes, as these may mar and destroy some of the bone surface and obliterate any traces of bone surface modifications such as cut and/or tooth marks. If water is used, bone should be dried slowly on a clean sheet of absorbent material out of direct heat or sunlight.
Never put wet bones in a plastic bag or any airtight container, as this can lead to mould and their eventual destruction. Bone should be stored in a dry place, in a clean, dry polyethylene bag into which small holes are made so that the bone does not ‘sweat’ in case any temperature fluctuations do occur. An acid-free tag containing any identifying information should be placed inside the bag with the bone. Writing on the outside of the bag is generally not recommended, as this can rub off and leave an unprovenanced bag of bone that is worthless.