2
Aboriginal plant use and ecological knowledge
47,000 years of monsoon rainforest connections in the Kimberley, north-west Australia
Introduction
Archaeobotany – the recovery and analysis of plant remains excavated from stratified archaeological contexts – provides insights into people’s diets and ecological relationships in the past. Taxonomic information obtained from archaeobotanical assemblages contributes greatly to reconstructions of diet, subsistence, landscape use, environmental conditions and palaeoclimate. A significant proportion of Australian Aboriginal diets and nutrition is met using food obtained from plants. However, the application of archaeobotany in Australian archaeology has been infrequent and the focus of archaeological discourse on lithic technologies and hunting strategies has largely neglected the role of plant foods and plant-related activities in Aboriginal societies, creating an incomplete picture of the past. This chapter begins with a brief overview of global archaeobotanical themes and Australian archaeobotany. It then focuses on the Kimberley region of Western Australia, where archaeobotanical research has revealed a rich and complex record of Aboriginal plant uses spanning 47,000 years of occupation.
Plants in the archaeological record
The visibility/invisibility of plants in the archaeological record has been a subject of ongoing inquiry since the first studies of plant remains derived from European archaeological contexts in the mid-nineteenth century (Hastorf 1999, 55). Visibility of plant remains is far less when compared to more robust cultural materials, such as stone tools (Hastorf 1999) from which it is only possible to infer plant use (e.g. Hayes et al. 2018; Hiscock et al. 2016; Maloney and Dilkes-Hall 2020; Wallis and O’Connor 1998). Direct evidence for food plants can be elusive and, in archaeological sites where plants do preserve, botanical remains represent only a fraction of what was originally used and deposited by humans; their occurrence in the archaeological record is often accidental (Yen 1988).
When preservation allows, archaeobotanical research can provide considerable information on people-plant relationships in the past and human interaction with the surrounding environment by answering questions about plant-based economies, subsistence strategies, cultural preferences, resource scheduling, seasonality, ecological targeting, habitat modification, mobility, land management strategies, domestication processes, agriculture, horticulture, arboriculture, environmental and climatic conditions, and, by extension, social ornamentation, identity, maritime capabilities and other plant-based perishable technologies (e.g. Antolín et al. 2016; Balme 2013; Balme and Morse 2006; Balme et al. 2022; Cappers and Neef 2012; Denham et al. 2009; Fuller 2018; Hastorf 1999; Hather 1994; Hather and Mason 2002; Pearsall 2010; Sayok and Teucher 2018).
Types of archaeobotanical remains
Plant remains enter archaeological sites via two main pathways: anthropogenic (direct and indirect) and non-anthropogenic introduction (Gallagher 2014, 29). It is important that culturally and naturally introduced archaeobotanical remains are differentiated as the two provide different types of information – the former reflective of diet, subsistence and human agency, the latter reflecting the environmental setting (Minnis 1981). Microbotanical remains include pollen, phytoliths and starch grains, while macrobotanical remains include wood charcoal, uncharred wood, bark, stems, leaves, flowers, fruits, seeds, nuts and modified botanical materials such as string, wooden tools and other plant-based technologies (Pearsall 2010).
This chapter is primarily concerned with macrobotanical remains and, more specifically, seed/fruit/nut remains, which – when shown to be introduced anthropogenically – offer information directly related to plants chosen for food by societies in the past.
Preservation of macrobotanical remains
All types of macrobotanical remains have the best potential to preserve in extreme environmental conditions, which prevent physical and chemical decay of organic plant materials and inhibit destructive biological processes (e.g. dry/wet/cold), or through exposure to fire, which transforms organic material to carbon (Miksicek 1987, 213–19). In Australian contexts, the two most common modes of preservation of macrobotanical remains are carbonisation and desiccation.
As with all organic archaeological remains, preservation and/or decomposition of plant material over time is complicated by several pre-depositional, depositional and post-depositional processes. Consequently, the quality, quantity and type of botanical material preserved can vary enormously within and across archaeological sites, producing intra- and inter-site difference/s (Clarke 1989; Hather 1994). Figure 2.1 traces various sources of botanical materials and how these enter archaeological sites alongside taphonomic processes that influence the probability of preservation.
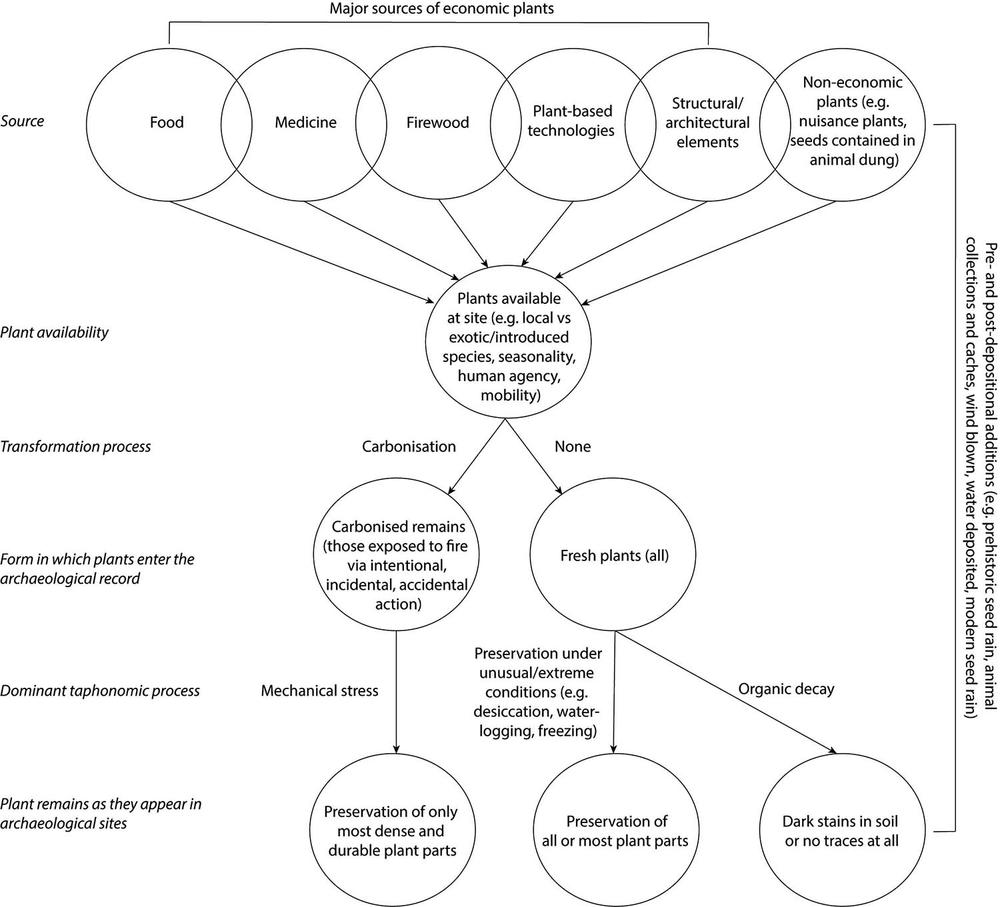
Figure 2.1 Anthropogenic and non-anthropogenic sources of macrobotanical remains and pre-depositional, depositional and post-depositional processes (adapted from Bush 2004, Figure 2.1, 19).
Global archaeobotanical themes
Globally, archaeobotanical investigation concentrates on origin centres of plant domestication and the rise and spread of agricultural societies (e.g. Denham 2018; Heiser 1990; Piperno 2018; Reed 1977; Ucko and Dimbleby 1969). Preoccupation of archaeobotanical studies with agriculture is underpinned by the fact that long-term human exploitation of botanical resources can have significant effects on plant morphology and genetics (Fuller and Allaby 2009), and morphological changes in plant structures associated with human manipulation can usually be identified and observed in the archaeobotanical record (Hancock 2004).
Changes to people-plant relationships in foraging societies are less visible because different plant propagation mechanisms (i.e. short-lived annuals versus long-lived perennials and vegetatively propagated crops) mean domestication of some plants is not always possible or morphologically distinguishable. In terms of morphological change in macrobotanical remains, domestication of perennial fruiting trees, shrubs and vines, and, to a lesser extent, underground storage organs, is not as well understood as it is for annual cereal crops (Denham et al. 2020; Fuller 2018).
Recent research has aimed to address the role of plants in foraging societies (e.g. Antolín et al. 2016; Hardy and Kubiak-Martens 2016; Mooney and Martín-Seijo 2021). However, the focus of archaeobotanical research on the origins of plant domestication and agricultural societies has perpetuated the notion that transitions from foraging to farming represent the only significant change in people-plant relationships in the past, presenting agriculture as an inevitable end on a linear trajectory (e.g. Ford 1985, 6; Harris 1990, 39). An example of the pervasiveness of the farming/foraging dichotomy can be observed in the traditional division of the Pleistocene continent Sahul (Australia, Tasmania, New Guinea and Aru Islands) into the Melanesian agricultural north and the Australian Aboriginal hunter-gatherer-fisher south (Figure 2.2).
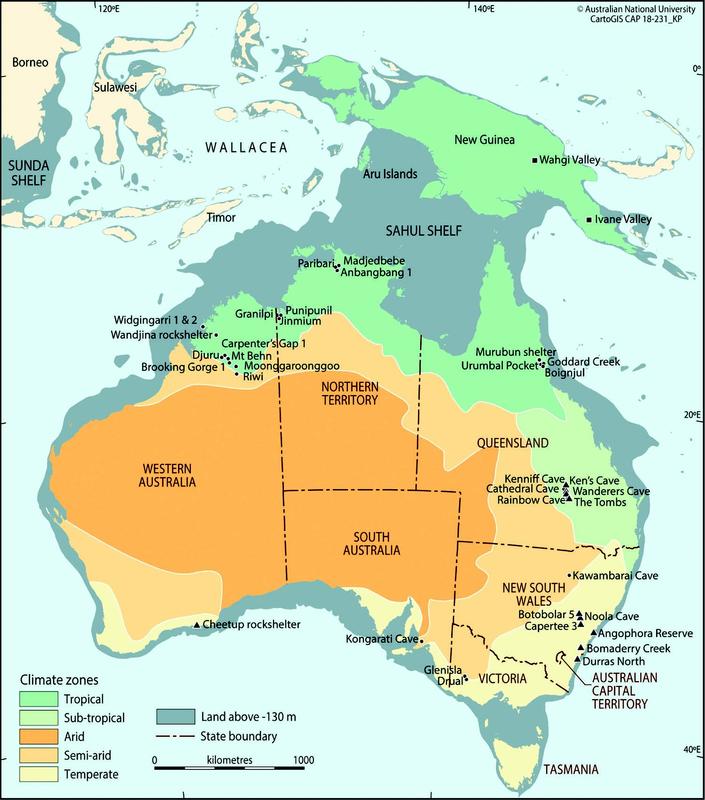
Figure 2.2 Australia with climatic zones, Last Glacial Maximum (LGM) coastline, and archaeological sites with non-woody seed/fruit macrobotanical investigation. Archaeological sites associated with plant domestication and agriculture display ■ icon. Archaeological sites associated with toxic plants display▲ icon (adapted from Whitau 2018, Figure 2.4, 46; CAD: CartoGIS, Australian National University).
Although efforts have been made to shake the divide (e.g. Florin and Carah 2018) and terms more inclusive of Australian Aboriginal plant uses and landscape management have been created and recreated over time, e.g. plant husbandry (Higgs and Jarman 1972), domiculture (Hynes and Chase 1982), incipient agriculture (Ford 1985), plant mothering (Hastorf 1998), low-level food production (Smith 2001), vegeculture (Barton and Denham 2018) and ecoscaping (Ouzman et al. 2019), archaeobotanical research in the region continues to concentrate on New Guinea as one of the global centres of plant domestication and agriculture (e.g. Denham 2018; Golson et al. 2017; Piperno 2018; Spengler III 2020). In contrast, less attention has been paid to archaeobotanical evidence from Australian contexts that can help to shed light on people-plant relationships and changes in plant use over time.
Australian archaeobotany
The late 1970s and 80s saw increased interest in archaeobotany by Australian researchers (e.g. Beaton 1977; Beck 1980; Clarke 1987; Head 1984) culminating in the seminal and unique monograph Plants in Australian Archaeology (Beck et al. 1989; see Brockwell et al. 2016 for a comprehensive overview of research on plants in Australian archaeology). Despite calls to shift Australian archaeobotanical research from the periphery to the core of archaeological pursuits (Denham et al. 2009), studies have remained sporadic. Overall, macrobotanical (here and hereafter excluding wood charcoal) research across the Australian continent is fragmentary, mainly targeting the eastern seaboard, and has largely centred on the use of toxic plants (e.g. Macrozamia). Figure 2.2 demonstrates macrobotanical remains preserved in different environments and site types across Australia. Despite this, “lack of preservation” is consistently cited by archaeologists who commonly draw upon this blanket statement to support their lack of engagement with archaeobotanical theory, method, fieldwork technique/s and research. This is especially astounding given that a considerable proportion (>50 per cent) of the dietary needs of Aboriginal peoples are met using plant foods (e.g. Kaberry 1935; McArthur 1960; Meehan 1989; Meggitt 1964; O’Dea et al. 1991; Russell-Smith et al. 1997), and the fact that plants are known to maintain important cultural connections to identity, language and Country (e.g. Blythe and Wightman 2003; Hercus 2012).
The publication of Dark Emu: Black Seeds: Agriculture or Accident? (Pascoe 2014) has been integral in drawing the attention of the Australian public and global readers to consider the complexities of Aboriginal plant use and resource management in the past. However, this has not taken place without significant conservative furore (O’Brien 2019, 2021a) and academic criticism (Keen 2021; Porr and Vivian-Williams 2021; Sutton and Walsh 2021). While it is not the purpose of this chapter to enter the debate surrounding Dark Emu, it is nonetheless important to highlight here that both Pascoe and critics alike fail entirely to consider and/or engage with available Australian archaeobotanical data (Australian examples include but are not limited to: Asmussen 2008, 2010; Asmussen and McInnes 2013; Atchison 2009; Atchison et al. 2005; Beck 1992; Byrne et al. 2021, 2019; Carah 2017; Clarke 1989, 1985; Cosgrove et al. 2007; Dotte-Sarout et al. 2015; Field et al. 2016; Ferrier and Cosgrove 2012; Florin et al. 2021, 2020; Hayes et al. 2018; King and Dotte-Sarout 2019; McConnell 1998; McConnell and O’Connor 1997; Owen et al. 2019; Roberts et al. 2021; Smith 1982; Walsh 2021; Walsh et al. 2024; Whitau et al. 2018a, 2018b, 2017).
Aboriginal ecological knowledge and monsoon rainforest connections spanning 47,000 years in the Kimberley
Study area
Recently, analyses of macrobotanical data from nine limestone cave and rock-shelter sites in the Kimberley region of north-west Australia have provided a detailed history of Aboriginal ecological knowledge relating to plant use spanning 47,000 years of occupation (Dilkes-Hall 2019; Dilkes-Hall et al. 2019a, 2019b, 2019c; Dilkes-Hall et al. 2020a, 2020b). The archaeological sites (Brooking Gorge 1, Carpenter’s Gap 1, Djuru, Moonggaroonggoo, Mount Behn, Riwi, Wandjina rock-shelter, and Widgingarri 1 and 2) are distributed from the north-east Kimberley coast to the south-central interior, across some 370 km; their locations positioning them across four Native Title determined lands: Dambimangari, Wilinggin, Bunuba, and the southernmost Gooniyandi (Figure 2.3).
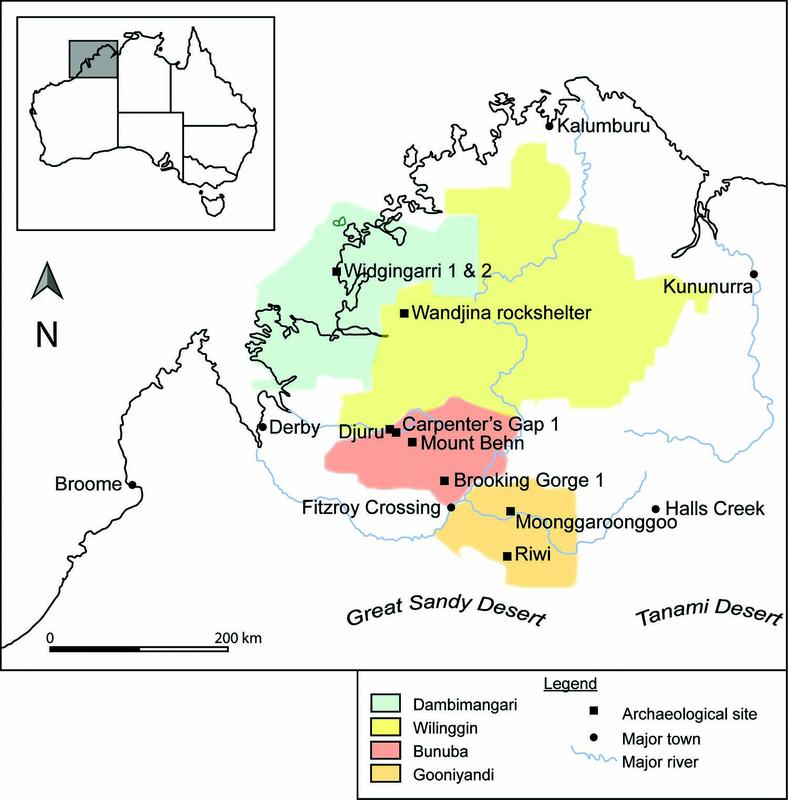
Figure 2.3 Map of the Kimberley showing locations of the archaeological sites with well-preserved macrobotanical sequences analysed for this research. Native Title determined lands after Kimberley Land Council (2022).
Monsoon rainforest
In the Kimberley, four major phytogeographic botanical districts – Dampier, Hall, Gardner and Fitzgerald after Beard (1979) – are overlapped by patches of monsoon rainforest (Figure 2.4). Monsoon rainforest occurs in coastal areas and persists further inland in fire protected gorges and limestone ranges and outcrops where water seepage maintains these sensitive vegetative communities (Beard 1976).
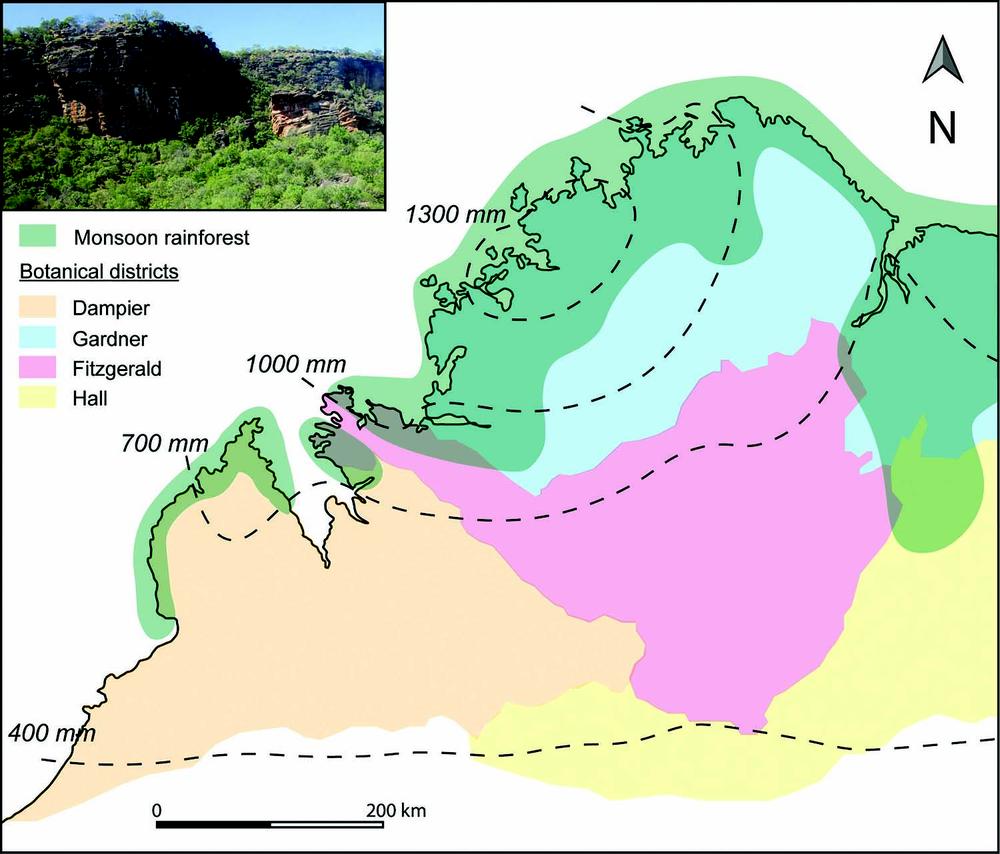
Figure 2.4 Map of the Kimberley region of north-west Western Australia. Botanical districts after Beard (1979). Present day monsoon rainforest distribution after Kenneally (2018). Rainfall isohyets after Bureau of Meteorology (1996).
The monsoonal climate of northern Western Australia strongly influences the seasonal availability of plant resources. Monsoon rainforests are floristically rich in plant species that – being physiologically adapted to monsoonal climatic conditions – rely on seasonal rainfall (November–April) for fruit development (Kenneally 2018). Plant species associated with monsoon rainforest are rare if not entirely absent from the surrounding open woodlands and include Celtis strychnoides (hackberry), Ficus spp. (fig), Flueggea virosa (white currant), Grewia spp. (currant bush), Terminalia spp. (terminalia) and Vitex spp. (black plum) (Kenneally 2018).
Preservation
At all nine of these Kimberley archaeological sites, macrobotanical materials were preserved primarily by desiccation. Generally, European convention favours analysis of only carbonised macrobotanical remains – fire as an inference for cultural association – discounting desiccated materials as modern in origin (Diestch 1996; Keepax 1997; Minnis 1981). Cultural aversion to burning particular taxa and/or specific plant parts in camp fires is not uncommon among Aboriginal groups in relation to food plants (Dilkes-Hall et al. 2019b); findings that are consistent with customs documented elsewhere in Australia concerning plants used for fuel (Byrne et al. 2019, 2013; Whitau et al. 2018a). Consequently, it is important to note here that carbonisation as a reliable analytical tool to separate anthropogenic from non-anthropogenic macrobotanical remains in Australian archaeological contexts is inconsistent with Aboriginal cultural practices and experiences. Therefore, the importance of dedicated analysis of desiccated components of macrobotanical assemblages cannot be overstated. As a result, this research analysed both desiccated and carbonised remains.
Methodology
Recovery techniques
Excavation and recovery techniques for each site have been described in detail elsewhere; Brooking Gorge 1, Djuru, Wandjina rock-shelter, and Widgingarri 1 and 2 (Dilkes-Hall et al. 2020b), Carpenter’s Gap 1 (Dilkes-Hall et al. 2019a), Moonggaroonggoo (Dilkes-Hall 2019), Mount Behn (Whitau et al. 2018) and Riwi (Dilkes-Hall et al. 2020a). For this research, working with previously excavated materials (legacy collections), difficulties accessing water during fieldwork in remote semi-arid/arid areas, and the fact that most macrobotanical materials are preserved by desiccation and so would be harmed by water exposure (Pearsall 2010, 80–1), meant flotation, which separates charred botanical material from sediment and recovers small botanical remains (Fairbairn 2005; Pearsall 2010), was not carried out for the majority of sites. All analysed material derive from dry sieving, except in the case of Mount Behn where flotation was used in conjunction with dry sieving (see Whitau et al. 2018).
Taxonomic identification
The first stages of taxonomic identification see macrobotanical remains grouped into analytical units based on morphological similarities, such as shape, dimension, length, width, surface and texture (see Fritz and Nesbitt 2014). Other attributes, such as dispersal mechanisms and rodent gnaw marks, were recorded to help determine non-anthropogenic taxa (Dilkes-Hall et al. 2019a, 37).
Grouped according to morphological attributes, taxonomic identifications are made by referring to one or more comparative reference collections of modern vouchered botanical specimens. Unavailability of comparative reference material can result in taxonomic misidentification generating misinterpretation (see Dilkes-Hall et al. 2019a). Development of collections is time-consuming, costly, often developed by individual researchers and/or for specific projects and are rarely readily accessible or made available digitally. Access to and development of reference collections is the single largest obstacle facing Australian archaeobotanical research today.
For this research, it was necessary to visit and document existing archaeobotanical and botanical collections housed in universities, museums and herbaria, and, in order to fill remaining gaps in the comparative reference collection, conduct botanical collection in the Kimberley region. The physical collection is housed in the archaeology laboratory at the University of Western Australia and is available to researchers. In conjunction, a database (FileMaker Pro) collates images and morphological descriptions of identified archaeobotanical remains alongside botanical information and examples of modern specimens, and is available on request.
Quantification
Quantification of macrobotanical remains included absolute counts (number of identified specimens [NISP]), mass, presence/absence and relative frequencies (these and other quantitative methods outlined in Popper 1988, 53–71).
To account for degrees of fragmentation disregarded by NISP, attempts were made to quantify minimum number of individuals (MNI). For this research, MNI was calculated by determining a single common characteristic distinct to each taxon. Although not generally accepted for publication, MNI is worthy of consideration to highlight variable patterns of fragmentation within and across taxa. For example, analysis of natural patterns of fragmentation of Vitex glabrata (Figure 2.5) allowed for the identification of accurate diagnostic elements with which to calculate MNI (e.g. the calyx, the whole fruit or a whole endocarp) and provided direct evidence to compare processed fruits against (discussed further below).
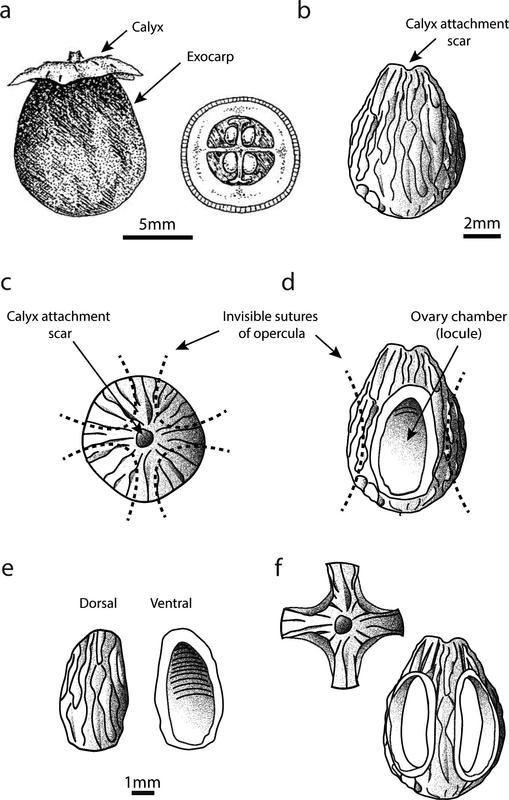
Figure 2.5 Vitex glabrata natural pattern of fragmentation: a) V. glabrata whole fruit profile and cross section (modified from Munir 1987, 45); b) Endocarp profile; c) Endocarp apex; d) Endocarp profile with operculum removed; e) Operculum; f) Endocarp structure with four opercula removed shown from the apex and profile.
Results
From the nine archaeological sites examined, a total of 21,833 macrobotanical remains were analysed (Table 2.1).
Fifty-seven taxa were identified to varying taxonomic levels (Dilkes-Hall
et al. 2020b, 1735). Forty-five of these are recognised as economically important plants by Aboriginal groups in the Kimberley today, contributing significantly to our understanding of the different economic, social and technological roles that plants played in Aboriginal lifeways in the past. Overall, taxonomic identification of macrobotanical remains demonstrates – consistently across time and space – one vegetation unit was primarily targeted for the collection of food plants in the Kimberley, monsoon rainforest.
| Archaeological site | NISP | Mass (g) |
|---|---|---|
| Brooking Gorge 1 | 4,138 | 151.39 |
| Carpenter’s Gap 1 | 7,846 | 185.56 |
| Djuru | 96 | 8.08 |
| Moonggaroonggoo | 604 | 22.14 |
| Mount Behn | 203 | 6.67 |
| Riwi | 7,609 | 61.18 |
| Wandjina rock-shelter | 487 | 5.82 |
| Widgingarri Shelter 1 | 739 | 74.79 |
| Widgingarri Shelter 2 | 111 | 14.34 |
| Total | 21,833 | 592.97 |
Table 2.1 Total NISP and mass of macrobotanical remains recovered from archaeological
sites.
Discussion
Seasonality, mobility and women
Seasonal movements of Aboriginal groups are evidenced in the macrobotanical archives because fruiting times of monsoon rainforest taxa correspond directly to periods of rainfall. As documented in recent history (Scarlett 1985), macrobotanical evidence shows the spatial location of Aboriginal campsites in the past is linked to the tempo of wet/dry climatic cycles of the monsoonal tropics (Dilkes-Hall et al. 2020b, 322). People’s movements to caves and rock-shelters during periods of rainfall have less to do with evading inclement weather and far more to do with ecological knowledge pertaining to resource availability, with people moving to locations where economic botanical resources are known to be coming into abundance (Dilkes-Hall et al. 2019b, 11).
A strong association exists between flora and fauna and, unsurprisingly, monsoon rainforest fruit production coincides with the collection of a number of important seasonal faunal resources (e.g. catfish, stingray, turtles, turtle eggs and goanna) because they too have reached their highest fat and nutritional content during this time of year (Crawford 1982; Davis et al. 2011; Smith and Kalotas 1985). Interestingly, these specific types of faunal resources are, as with plants, most often collected by women (Crawford 1982, 18; Davis, personal communication, 2016; O’Dea et al. 1991, 234). In this way, macrobotanical data available for the Kimberley region shows intimate connections between plants, animals, women, seasonality, landscape use and food traditions, which together strongly influence the configuration of social identity and group membership in Aboriginal societies by showing “that you belong to the country, that you are a product of the country” (Blythe and Wightman 2003, 69).
Plant processing and foodways
When discernible in macrobotanical archives, plant processing activities can provide important insights into processing techniques, cultural preferences and foodways in the past. At Riwi, numerous macrobotanical remains were documented as economic plants by Gooniyandi Traditional Owners (Figure 2.6) and food plants are dominated by monsoon rainforest species, in particular Vitex cf. glabrata (Gooniyandi name: girndi). Careful analysis of desiccated girndi remains, as opposed to discounting them altogether (e.g. Keepax 1977; Minnis 1981), provided extraordinary evidence of fruit processing (Dilkes-Hall et al. 2019b).

Figure 2.6 Macrobotanical remains recovered from Riwi documented as economic plants by Gooniyandi Traditional Owners: a) Acacia sp. Type A pod; b) Celtis strychnoides endocarps; c) Eucalyptus-Corymbia gall; d) Eucalyptus-Corymbia capsule; e) Ficus spp. fruits; f) Flueggea virosa seeds; g) Melaleuca spp. paperbark; h) Premna acuminata endocarp; i) Senna sp. seed; j) Terminalia sp. Type A (cf. ferdinandiana) endocarp; k) Triodia cf. pungens spikelets; l) Vitex cf. glabrata endocarps (from Dilkes-Hall et al. 2019c, Figure 7, 19).
Preservation of girndi calyces by desiccation is important to note here, not only acting as an accurate diagnostic element with which to calculate MNI (n=720), but providing evidence for stage one processing where whole fruits are dehydrated by the camp fire – a process that detaches calyces from fruits and makes robust endocarps brittle and easier to process during stage two (Dilkes-Hall et al. 2019b, 6). Fragmentation patterns produced by stage two processing were discernible from natural breakage patterns, with fragmentation as a result of processing, thereby producing jagged pieces that do not follow natural sutures (Figure 2.5) and cut across the strongest part of the endocarp, the operculum (Figure 2.7). At Riwi, fruit processing, food conservation and associated storage techniques dating to the mid Holocene indicate innovative socio-economic technological change to incorporate, manage and conserve seasonally abundant girndi fruits (Dilkes-Hall et al. 2019b).

Figure 2.7 Girndi (Vitex glabrata) from harvest to storage. Results shown across the three lines of evidence documented 1) ethnobotany 2) experimental archaeology 3) macrobotanical signature. Images produced by Dilkes-Hall using a Canon IXUS 180 digital camera and Dino-Lite Edge digital microscope (from Dilkes-Hall et al. 2019b, Figures 4, 9).
Here, macrobotanical evidence coupled with documentation of traditional ecological knowledge, plant collection and experimental archaeology worked together to shed light on women’s activities, introducing women into an archaeological narrative that they have largely been excluded from (e.g. Bowdler and Balme 2010;
Hastorf 1998; Watson and Kennedy 1991). Most importantly, this aspect of the research highlights and strengthens the cultural links between Gooniyandi women today, who maintain the tradition of collection and processing activities, and their ancestors who performed similarly crucial tasks and societal roles several thousand years ago.
Human responses to climate/environmental change
Macrobotanical research offers a unique opportunity to investigate questions around human responses to major palaeoclimatic and palaeoenvironmental changes recorded for the Kimberley since human occupation. Specifically, structural changes in vegetation are indicated by periods of peak aridity, such as the LGM (~18,000 years ago) and late Holocene El Niño–Southern Oscillation (ENSO) driven climate change (~4,200 years ago onwards) (Denniston et al. 2013; Lambeck et al. 2002). Surprisingly, during these sustained periods of aridity – when significant reorganisation of diet and subsistence in response to environmental change might be expected – continued use of monsoon rainforest taxa was observed in the macrobotanical archives with little evidence of dramatic changes to diet and subsistence (Dilkes-Hall et al. 2020b, 1736).
These findings are in contrast to changes indicated by other archaeological materials from some of the same sites under investigation, such as lithic and faunal records at Carpenter’s Gap 1 (Maloney et al. 2018), which provide evidence for adjustments coincident to climate and environmental changes. Ultimately, the results demonstrate that changes in people’s subsistence strategies and responses to environmental change vary across different economic resources (e.g. stone, fauna and flora), suggesting differences in women’s and men’s responses to climate change that may have affected the activities and roles of these gendered groups differently, and/or that monsoon rainforest – especially escarpment forests – remained steady as a resource zone despite climate change (Russell-Smith 1985, 243).
A common botanical heritage, colonisation and ecological knowledge
Overall, macrobotanical evidence from the Kimberley indicates a botanical heritage shared by Indigenous peoples across the Indo-Pacific region and other parts of the world. For example, Terminalia species have also been recovered from archaeological sites in Papua New Guinea (Gorecki et al. 1991) and Timor-Leste (Oliveria 2008). Adansonia, Grewia and Vitex species have been identified in African macrobotanical records (Kahlheber et al. 2009; Neumann et al. 1998; Sievers 2006). Celtis species have been recovered from archaeological sites located in Africa (Kahlheber et al. 2009; Sievers 2006), Georgia (Messager et al. 2010), Israel (Simchoni et al. 2011), New Ireland (Rosenfeld 1997), Papua New Guinea (Fredericksen et al. 1993) and Turkey (Fairbairn et al. 2002). Canarium species are present in macrobotanical records from sites in Africa (Kahlheber et al. 2009), Sri Lanka (Perera et al. 2011; Wedage et al. 2020), Borneo (Barton et al. 2016; Dilkes-Hall et al. forthcoming), the Philippines (Pawlik et al. 2014) and New Guinea and Near and Remote Oceania (Fairbairn and Florin 2022). This interconnectedness indicates a common botanical heritage with foundations along migration pathways to Sahul.
Golson (1971, 209) hypothesised that continental colonisation by Aboriginal groups was assisted by familiarity with Indo-Malaysian plants found in monsoon rainforests across northern Australia. The presence of important Indo-Malaysian botanical elements, Terminalia and Vitex species, in the earliest cultural unit at Carpenter’s Gap 1, dating to 51–39 ka (Dilkes-Hall et al. 2019a, 34), is testimony to this. Early colonising groups likely encountered Terminalia and Vitex species throughout Sunda and Island Southeast Asia and entered northern Sahul equipped with ecological knowledge of nutritious fruit-bearing trees of these genera, successfully applying this taxonomy to monsoon rainforest environments as observed at Carpenter’s Gap 1 (Dilkes-Hall et al. 2019a) and earlier still in the Northern Territory at Madjedbebe by 65–53 ka (Florin et al. 2020).
Transmission of ecological knowledge onto new landscapes likely facilitated expedient identification of nutritious monsoon rainforest plants, easing the process of having to adapt to otherwise totally unknown environments. Available macrobotanical data for the Kimberley indicates monsoon rainforests represented secure and familiar environments to Aboriginal groups, demonstrating their importance in Aboriginal foodways and subsistence systems over 47,000 years of occupation.
Despite dramatic alteration to Aboriginal lifeways since European invasion, monsoon rainforest ecosystems remain a vital resource for Aboriginal groups today. Monsoon rainforests are often associated with important cultural sites and are actively protected from wildfires using traditional land management practices, particularly controlled fire, to maintain boundaries between rainforest and savannah (Vigilante et al. 2017). Maintenance of these ecosystems goes far beyond the simple protection of important economic plant species with these deeply socio-cultural activities encompassing affirmations of group identity, kinship systems, language and cultural connections to Country.
Conclusions
First Nations engagement and archaeobotanical research
To explore Aboriginal plant uses successfully using archaeobotanical evidence, it is fundamental that researchers work closely with Traditional Owners – the local experts in both contemporary and traditional ecological knowledge. Partnership and collaboration with Traditional Owners is essential to understanding plant use in the past and, through two-way learning, mutual benefits can be gained. For example, plant use can be documented with Traditional Owners on botanical surveys and collection, and researchers can demonstrate analytical techniques, return archaeological information pertaining to past plant use, develop interpretations with Traditional Owners and support First Nations authorship.
Engagement between researchers and Traditional Owners has great benefits for knowledge and understanding of plant use, plant resource management, women’s activities, landscape use, changing subsistence strategies and responses to environmental change. Collaboration and engagement at the earliest stages of planning shapes research projects for the better (e.g. Maloney et al. 2017) and educates researchers on how archaeological and archaeobotanical records might be better observed and interpreted in a way that has meaningful outcomes for local Aboriginal groups (Davis et al. 2021; Dilkes-Hall et al. 2019c).
Resilience through continuity
Overall, from the macrobotanical evidence available for the Kimberley region, a broad picture of continuity emerges. Here, the notion of continuity should not be taken to suggest a static continuum of botanical knowledge. Instead, macrobotanical evidence suggests that complex socio-economic strategies, such as seasonal scheduling, fruit processing technologies, and landscape management practices (e.g. fire regimes and translocation of economic botanical species), were likely to have been employed by Aboriginal groups to ensure the future availability of important botanical resources (fruit-bearing monsoon rainforest trees) across not only time – as evidenced in the macrobotanical archives by the continued use of these food plants – but also geographical space – as evidenced by the spatial locations of archaeological sites analysed.
Continuity in plants targeted for food through time demonstrates the importance of resilience in both monsoon rainforest vegetation and the subsistence systems employed by Aboriginal groups that target this specific type of vegetation. Simultaneously, this research draws archaeological attention to the significant role of women in Aboriginal economies. Further, collection of plant foods, often carried out as a group activity, encompasses intangible aspects of social life, such as reinforcing identity and group cohesion and fulfilling the vital role of passing on important ecological knowledge to younger generations.
Since colonisation of Sahul ~65,000 years ago (Clarkson et al. 2017), persistent use of monsoon rainforest fruits through to the present demonstrates botanical knowledge carried by early Aboriginal populations was passed down intergenerationally. In the Kimberley, macrobotanical evidence shows transmission of ecological knowledge over 47,000 years highlighting the important role of Aboriginal oral traditions and the essential function/s that these perform in culture, society and economy while informing our understanding of vulnerability and resilience to climate change in the past.
Acknowledgements
The author India Ella Dilkes-Hall (IEDH) thanks the Dambimangari, Wilinggin, Bunuba and Gooniyandi Aboriginal communities of the lands where archaeological excavations took place. Specifically, IEDH acknowledges support from Gooniyandi Traditional Owners, Muludja community members and Gooniyandi Rangers. A special thanks to Rosemary Nuggett and the Mimbi Community. To record ethnobotanical knowledge with Gooniyandi Traditional Owners, ethics approval was granted by the Human Ethics Office at the University of Western Australia (RA/4/1/8255). The Girndi Project, undertaken with senior Gooniyandi Traditional Owners June Davis and Helen Malo, was supported by Graduate Women (WA) Jillian Bradshaw Scholarship 2017 and the Australian Archaeological Association Student Research Grant Scheme 2017 received by IEDH. Thank you to Matthew David Barrett, Russell Lindsey Barrett and Kevin Kenneally for assistance with taxonomic identifications over the course of my doctoral research. At time of writing, IEDH was a Forrest Foundation Prospect Fellow supported by the Forrest Research Foundation. In August 2018, June Davis† passed away. Her laughter sings out across Gooniyandi Country for eternity. Her family and community have asked that her name and photos continue to be used in recognition of her great contributions to the research. A special mention to Dr Tim Ryan Maloney†, without whom crucial parts of this research would not have been possible. Simply the best.
References
Antolín, F., M.B. Azorin and O. López-Bultó (2016). Archaeobotany of wild plant use: approaches to the exploitation of wild plant resources in the past and its social implications. Quaternary International 404: 1–3.
Asmussen, B. (2010). In a nutshell: the identification and archaeological application of experimentally defined correlates of Macrozamia seed processing. Journal of Archaeological Science 37(9): 2117–25.
Asmussen, B. (2008). Anything more than a picnic? Re-considering arguments for ceremonial Macrozamia use in mid-Holocene Australia. Archaeology in Oceania 43(3): 93–103.
Asmussen, B. and P. McInnes (2013). Assessing the impact of mid-to-late Holocene ENSO-driven climate change on toxic Macrozamia seed use: a 5,000 year record from eastern Australia. Journal of Archaeological Science 40(1): 471–80.
Atchison, J. (2009). Human impacts on Persoonia falcata. Perspectives on post-contact vegetation change in the Keep River region, Australia, from contemporary vegetation surveys. Vegetation History and Archaeobotany 18(2): 147–57.
Atchison, J., L. Head and R. Fullagar (2005). Archaeobotany of fruit seed processing in a monsoon savanna environment: evidence from the Keep River region, Northern Territory, Australia. Journal of Archaeological Science 32(2): 167–81.
Balme, J. (2013). Of boats and string: the maritime colonisation of Australia. Quaternary International 285: 68–75.
Balme, J. and K. Morse (2006). Shell beads and social behaviour in Pleistocene Australia. Antiquity 80(310): 799–811.
Balme, J., S. O’Connor, T.R. Maloney, K. Akerman, B. Keaney and I.E. Dilkes-Hall (2022). Fibre technologies in Indigenous Australia: evidence from archaeological excavations in the Kimberley region. Australian Archaeology 88(2): 115–28. DOI: 10.1080/03122417.2022.2054510.
Barton, H. and T. Denham (2018). Vegecultures and the social-biological transformations of plants and people. Quaternary International 489: 17–25.
Barton, H., V. Paz and A.J. Carlos (2016). Plant food remains from the Niah caves: macroscopic and microscopic approaches. In G. Barker and L. Farr, eds. Archaeological investigations in the Niah caves, Sarawak, The Archaeology of the Niah Caves, Sarawak, vol. 2, 455–68. Cambridge: McDonald Institute for Archaeological Research.
Beard, J.S. (1979). The vegetation of the Kimberley area: explanatory notes to sheet 1. Perth: University of Western Australia Press.
Beard, J.S. (1976). The monsoon forests of the Admiralty Gulf, Western Australia. Vegetatio 31(3): 177–92.
Beaton, J.M. (1977). Dangerous harvest: investigations in late pre-historic occupation of upland south-east central Queensland. Doctoral thesis, Australian National University, Canberra, ACT.
Beck, W. (1992). Aboriginal preparation of Cycas seeds in Australia. Economic Botany 46(2): 133–47.
Beck, W. (1980). Aspects of plant taphonomy in Australian archaeology. Master’s thesis, La Trobe University, Melbourne, Vic.
Beck, W., A. Clarke and L. Head, eds (1989). Plants in Australian archaeology. TEMPUS, Series No. 1. St Lucia: Anthropology Museum, University of Queensland.
Blythe, J. and G. Wightman (2003). The role of animals and plants in maintaining links. In J. Blythe and R.M. Brown, eds. Maintaining the links: language, identity and the land. Proceedings of the seventh conference presented by the Foundation for Endangered Languages, 69–77. Bath: Foundation for Endangered Languages.
Bowdler, S. and J. Balme (2010). Gatherers and grannies: further thoughts on the origins of gender. Australian Feminist Studies 25(66): 391–405.
Brockwell, S., J. Stevenson and A. Clarke (2016). Plants and archaeology in Australia. In K. Hardy and L. Kubiak-Martens, eds. Wild harvest: plants in the hominin and pre-agrarian human worlds, 273–99. Oxford: Oxbow Books.
Bureau of Meteorology (1996). Kimberley, Western Australia: climatic survey. Canberra: Australian Government Publishing Service.
Byrne, C., E. Dotte-Sarout and V. Winton (2013). Charcoals as indicators of ancient tree and fuel strategies: an application of anthracology in the Australian midwest. Australian Archaeology 77(1): 94–106.
Byrne, C., T. Dooley, T. Manne, A. Paterson and E. Dotte-Sarout (2019). Island survival: the anthracological and archaeofaunal evidence for colonial-era events on Barrow Island, north-west Australia. Archaeology in Oceania 55(1): 15–32.
Byrne, C., E. Dotte-Sarout, S. van Leeuwen, J. McDonald and P. Veth (2021). The dependable deep time Acacia: anthracological analysis from Australia’s oldest Western Desert site. Journal of Archaeological Science: Reports 40: 103–87.
Bush, L.L. (2004). Boundary conditions: macrobotanical remains and the Oliver Phase of central Indiana, A.D. 1200–1450. Tuscaloosa: University of Alabama Press.
Cappers, R.T.J. and R. Neef (2012). Handbook of plant palaeoecology. Groningen: Barkhuis.
Carah, X.A. (2017). Regime change: an anthracological assessment of fuel selection and management at Madjedbebe (Malakunanja II), Mirarr country, Australia. Doctoral thesis, University of Queensland, Brisbane, Qld.
Clarke, A. (1989). Macroscopic plant remains. In W. Beck, A. Clarke and L. Head, eds. Plants in Australian archaeology, 54–89. Tempus, Series No. 1. St Lucia: Anthropology Museum, University of Queensland.
Clarke, A. (1987). An analysis of archaeobotanical data from two sites in Kakadu National Park, Northern Territory. Master’s thesis, University of Western Australia, Crawley, WA.
Clarke, A. (1985). A preliminary archaeobotanical analysis of the Anbangbang I site. In R. Jones, ed. Archaeological research in Kakadu National Park, 77–96. Australian National Parks and Wildlife Service, Special Publication 13. Canberra: Australian National University.
Clarkson, C., Z. Jacobs, B. Marwick, R. Fullagar, L.A. Wallis, M. Smith et al. (2017). Human occupation of northern Australia by 65,000 years ago. Nature 547(7663): 306–13.
Cosgrove, R., J. Field and A. Ferrier (2007). The archaeology of Australia’s tropical rainforests. Palaeogeography, Palaeoclimatology, Palaeoecology 251(1): 150–73.
Crawford, I.M. (1982). Traditional Aboriginal plant resources in the Kalumburu area: aspects in ethno-economics. Perth: Western Australian Museum.
Davis, J., M. Street, H. Malo, I. Cherel and E. Woodward (2011). Mingayooroo – Manyi Waranggiri Yarrangi. Gooniyandi seasons (calendar) Margaret River, Fitzroy Valley, Western Australia. Darwin: CSIRO Ecosystem Sciences.
Davis, J., H. Malo, E. Cherel, M. Street, W. Cherrabun, B. Cherel and I.E. Dilkes-Hall (2021). Gooniyandi Binarri. Crawley: The Author.
Denham, T.P. (2018). Tracing early agriculture in the highlands of New Guinea: plot, mound and ditch. Oxford: Routledge.
Denham, T., J. Atchison, J. Austin, S. Bestel, D. Bowdery, A. Crowther, N. Dolby, A. Fairbairn, J. Field and A. Kennedy (2009). Archaeobotany in Australia and New Guinea: practice, potential and prospects. Australian Archaeology 68(1): 1–10.
Denham, T., H. Barton, C.C. Castillo, A. Crowther, E. Dotte-Sarout, S.A. Florin, J. Pritchard, A. Barron, Y. Zhang and D.Q. Fuller (2020). The domestication syndrome in vegetatively propagated field crops. Annals of Botany 125: 581–97.
Denniston, R.F., K.H. Wyrwoll, V.J. Polyak, J.R. Brown, Y. Asmerom, A.D. Wanamaker Jr et al. (2013). A stalagmite record for Holocene Indonesian-Australian summer monsoon variability from the Australian tropics. Quaternary Science Reviews 78: 155–68.
Dietsch, M-F. (1996). Gathered fruits and cultivated plants at Bercy (Paris), a Neolithic village in a fluvial context. Vegetation History and Archaeobotany 5(1/2): 89–97.
Dilkes-Hall, I.E. (2019). Macrobotanical remains and preservational bias: an example from Moonggaroonggoo, Gooniyandi Country, Western Australia. Australian Archaeology 85(2): 210–14.
Dilkes-Hall, I.E., S. O’Connor and J. Balme (2019a). People-plant interaction and economic botany over 47,000 years of occupation at Carpenter’s Gap 1, south central Kimberley. Australian Archaeology 85(1): 30–47.
Dilkes-Hall, I.E., J. Davis and H. Malo (2019b). “Doog girndi”. Using experimental archaeology to understand the archaeobotanical record: an investigation of mid-Holocene Vitex glabrata fruit processing in Gooniyandi Country, northwest Australia. The Artefact 42: 3–16.
Dilkes-Hall, I.E., T.R. Maloney, J. Davis, H. Malo, E. Cherel, M. Street,
W. Cherrabun and B. Cherel (2019c). Understanding archaeobotany through ethnobotany: an example from Gooniyandi Country, northwest, Western Australia. Journal of Anthropological Society of South Australia 43: 5–33.
Dilkes-Hall, I.E., J. Balme, S. O’Connor and E. Dotte-Sarout (2020a). Archaeobotany of Aboriginal plant foods during the Holocene at Riwi, south central Kimberley, Western Australia. Vegetation History and Archaeobotany 29(3): 309–25.
Dilkes-Hall I.E., J. Balme, S. O’Connor and E. Dotte-Sarout (2020b). Evaluating people’s responses to ENSO driven climate change during the Holocene in northwest Australia through macrobotanical analyses. The Holocene 30 (12): 1728–40.
Dilkes-Hall, I.E., T.R. Maloney, E. Sriputri, A. Priyatno, Febryanto, M. Effendy et al. (forthcoming). Planting the seeds: an overview of archaeobotany in Borneo and new directions for East Kalimantan. KALPATARU.
Dotte-Sarout, E., X. Carah and C. Byrne (2015). Not just carbon: assessment and prospects for the application of anthracology in Oceania. Archaeology in Oceania 50(1): 1–22.
Fairbairn, A.S. and S.A. Florin (2022). Archaeological identification of fragmented nuts and fruits from key Asia-Pacific economic tree species using anatomical criteria: comparative analysis of Canarium, Pandanus and Terminalia. Archaeology in Oceania 57: 160–88.
Fairbairn, A., E. Asouti, J. Near and D. Martinoli (2002). Macro-botanical evidence for plant use at neolithic Çatalhöyük, south-central Anatolia. Turkey. Vegetation History and Archaeobotany 11(1): 41–54.
Ferrier, A. and R. Cosgrove (2012). Aboriginal exploitation of toxic nuts as a late-Holocene subsistence strategy in Australia’s tropical rainforests. In S. Haberle and B. David, eds. Peopled landscapes: archaeological and biogeographical approaches to landscapes, 103–20. Terra Australis 34. Acton: Australian National University E Press.
Field, J.H., L. Kealhofer, R. Cosgrove and A.C.F. Coster (2016). Human-environment dynamics during the Holocene in the Australian wet tropics of NE Queensland: a starch and phytolith study. Journal of Anthropological Archaeology 44: 216–34.
Florin, S.A. and X. Carah (2018). Moving past the “Neolithic problem”: the development and interaction of subsistence systems across northern Sahul. Quaternary International 489: 46–62.
Florin, S.A., A.S. Fairbairn, M. Nango, D. Djandjomerr, B. Marwick, R. Fullagar et al. (2020). The first Australian plant foods at Madjedbebe, 65,000–53,000 years ago. Nature Communications 11(924). DOI: 10.1038/s41467-020-14723-0.
Florin, S.A., P. Roberts, B. Marwick, N.R. Patton, J. Shulmeister, C.E. Lovelock et al. (2021). Pandanus nutshell generates a palaeoprecipitation record for human occupation at Madjedbebe, northern Australia. Nature Ecology and Evolution 5(3): 295–303.
Ford, R.I. (1985). Patterns of prehistoric food production in North America. In R.I. Ford, ed. Prehistoric food production in North America, 341–64. Michigan: University of Michigan, Museum of Anthropology.
Fredericksen, C., M. Spriggs and W. Ambrose (1993). Pamwak rockshelter:
a Pleistocene site on Manus Island, Papua New Guinea. In M.A. Smith,
M. Spriggs and B. Frankhauser, eds. Sahul in review: Pleistocene archaeology in Australia, New Guinea and Island Melanesia. Occasional papers in prehistory, No. 24, 144–52. Canberra: Australian National University.
Fritz, G. and M. Nesbitt (2014). Laboratory analysis and identification of plant macroremains. In J.M. Marston, J. D’alpoim Guedes and C. Warinner, eds. Method and theory in paleoethnobotany, 115–45. Boulder: University Press of Colorado.
Fuller, D.Q. (2018). Long and attenuated: comparative trends in the domestication of tree fruits. Vegetation History and Archaeobotany 27(1): 165–76.
Fuller, D.Q. and R. Allaby (2009). Seed dispersal and crop domestication: shattering, germination, and seasonality in evolution under cultivation. In L. Østergaard, ed. Fruit development and seed dispersal, 238–95. Annual Plant Reviews Volume 38. Chichester: Blackwell Publishing.
Gallagher, D.E. (2014). Formation processes of the macrobotanical record. In J.M. Marston, J. D’alpoim Guedes and C. Warinner, eds. Method and theory in paleoethnobotany, 19–34. Boulder: University Press of Colorado.
Golson, J. (1971). Australian Aboriginal food plants: some ecological and culture-historical implications. In D.J. Mulvaney and J. Golson, eds. Aboriginal man and environment in Australia, 196–238. Canberra: Australian National University.
Golson, J., T.P. Denham, P.J. Hughes, P. Swadling and J. Muke, eds (2017). Ten thousand years of cultivation at Kuk Swamp in the highlands of Papua New Guinea. Terra Australis 46. Canberra: Australian National University E Press.
Gorecki, P., M. Mabin and J. Campbell (1991). Archaeology and geomorphology of the Vanimo coast, Papua New Guinea: preliminary results. Archaeology in Oceania 26(3): 119–22.
Hancock, J.F. (2004). Plant evolution and the origin of crop species. Wallingford: CABI Publishing.
Hardy, K. and L. Kubiak-Martens, eds (2016). Wild harvest: plants in the hominin and pre-agrarian human worlds. Oxford: Oxbow Books.
Harris, D.R. (1990). Settling down and breaking ground: rethinking the Neolithic revolution. Amsterdam: Stichting Nederlands Museum voor Anthropologie en Prehistorie, Twaalfde Kroon-Voordracht.
Hastorf, C.A. (1999). Recent research in paleothnobotany. Journal of Archaeological Research 7(1): 55–103.
Hastorf, C.A. (1998). The cultural life of early domestic plant use. Antiquity 72(278): 773–82.
Hather, J.G. (1994). Tropical archaeobotany: applications and new developments. London: Routledge.
Hather, J.G. and S.L.R. Mason, eds (2002). Hunter-gatherer archaeobotany: perspectives from the northern temperate zone. London: Institute of Archaeology, University College London.
Hayes, E., R. Fullagar, K. Mulvaney and K. Connell (2018). Food or fibercraft? Grinding stones and Aboriginal use of Triodia grass (spinifex). Quaternary International 468(Part B): 271–83.
Head, L. (1984). Environment as artefact: a palaeoecological contribution to the prehistory of southwestern Victoria. Doctoral thesis, Monash University, Melbourne, Vic.
Heiser, C.B. (1990). Seed to civilization. Cambridge: Harvard University Press.
Hercus, L. (2012). Trees from the Dreaming. The Artefact 35: 40–9.
Higgs, E. and M.R. Jarman (1972). The origins of animal and plant husbandry. In E. Higgs, ed. Papers in economic prehistory, 3–13. Cambridge: Cambridge University Press.
Hiscock, P., S. O’Connor, J. Balme and T. Maloney (2016). World’s earliest ground-edge axe production coincides with human colonisation of Australia. Australian Archaeology 82(1): 2–11.
Hynes, R.A. and A.K. Chase (1982). Plants, sites and domiculture: Aboriginal influence upon plant communities in Cape York Peninsula. Archaeology in Oceania 17(1): 38–50.
Kaberry, P. (1935). The Forrest River and Lyne River tribes of north-west Australia: a report on field work. Oceania 5(4): 408–36.
Kahlheber, S., A. Höhn and N. Rupp (2009). Archaeobotanical studies at Nok sites: an interim report. Nyame Akuma 71: 2–17.
Kenneally, K.F. (2018). Kimberley tropical monsoon rainforest of Western Australia: perspectives on biological diversity. Journal of the Botanical Research Institute of Texas 12(1): 149–228.
Keen, I. (2021). Foragers or farmers: dark emu and the controversy over Aboriginal agriculture. Anthropological Forum 31(1): 106–28.
Keepax, C. (1977). Contamination of archaeological deposits by seeds of modern origin with particular reference to the use of flotation machines. Journal of Archaeological Science 4(3): 221–9.
King, F. and E. Dotte-Sarout (2019). Wood charcoal analysis in tropical rainforest: a pilot study identifying firewood used at toxic nut processing sites in northeast Queensland, Australia. Vegetation History and Archaeobotany 28(2): 163–85.
Kimberley Land Council (2022). Native title map, KLC website, accessed 13 January 2022. https://www.klc.org.au/native-title-map.
Lambeck, K., Y. Yokoyama and T. Purcell (2002). Into and out of the Last Glacial Maximum: sea-level change during Oxygen Isotope Stages 3 and 2. Quaternary Science Reviews 21: 343–360.
Maloney, T.R. and I.E. Dilkes-Hall (2020). Assessing the spread and uptake of tula adze technology in the late Holocene across the southern Kimberley of Western Australia. Australian Archaeology 86(3): 264–83.
Maloney, T., I.E. Dilkes-Hall and J. Davis (2017). Indigenous led archaeological excavation at Moonggaroonggoo, Gooniyandi country, Western Australia, reveals late Holocene occupation. Australian Archaeology 83(3): 178–84.
Maloney, T., S. O’Connor, R. Wood and J. Balme (2018). Carpenters Gap 1: A 47,000 year old record of Indigenous adaption and innovation. Quaternary Science Reviews 191: 204–28.
McArthur, M. (1960). Food consumption and dietary levels of groups of Aborigines living on naturally occuring foods. In C.P. Mountford, ed. Record of the American-Australian expedition in Arnhem Land, Vol II, Anthropology and Nutrition, 90–135. Melbourne: University of Melbourne.
McConnell, K. (1998). The prehistoric use of Chenopodiaceae in Australia: evidence from Carpenter’s Gap Shelter 1 in the Kimberley, Australia. Vegetation History and Archaeobotany 7(3): 179–88.
McConnell, K. and S. O’Connor (1997). 40,000 year record of food plants in the southern Kimberley ranges, Western Australia. Australian Archaeology 45(1): 20–31.
Meehan, B. (1989). Plant use in a contemporary Aboriginal community and prehistoric implications. In W. Beck, A. Clarke and L. Head, eds. Plants in Australian Archaeology, 14–30. Tempus, Series No. 1. St Lucia: Anthropology Museum, University of Queensland.
Meggitt, M.J. (1964). Aboriginal food-gatherers of tropical Australia. In The Ecology of Man in the tropical Environment, 30–37. International Union for Conservation of Native and Natural Resources, Ninth Technical Meeting, Publication Series 4. Morges, Switzerland: International Union for Conservation of Native and Natural Resources.
Messager, E., A. Badou, F. Fröhlich, B. Deniaux, D. Lordkipanidze and P. Voinchet (2010). Fruit and seed biomineralization and its effect on preservation. Archaeological and Anthropological Sciences 2(1): 25–34.
Miksicek, C.H. (1987). Formation processes of the archaeobotanical record. Advances in Archaeological Method and Theory 10: 211–47.
Minnis, P.E. (1981). Seeds in archaeological sites: sources and some interpretive problems. American Antiquity 46(1): 143–52.
Mooney, D.E. and M. Martín-Seijo (2021). Editorial: archaeobotany in the wider landscape. Environmental Archaeology 26(2): 115–21.
Munir, A.A. (1987). A taxonomic revision of the genus Vitex L. (Verbenaceae) in Australia Journal of the Adelaide Botanic Gardens 10(1): 31–79.
O’Brien, P. (2019). The bogus Aboriginal world of Bruce Pascoe. Quandrant online. https://quadrant.org.au/magazine/2019/12/the-bogus-aboriginal-world-of-brucepascoe/.
O’Brien, P. (2021a). A left-flank dismissal of “Dark Emu”. Quadrant online. quadrant.org.au/opinion/review/2021/07/a-left-flank-dismissal-of-dark-emu/.
Neumann, K., S. Kahlheber and D. Uebel (1998). Remains of woody plants from Saouga, a medieval west African village. Vegetation History and Archaeobotany 7(2): 57–77.
O’Dea, K., P.A. Jewell, A. Whiten, S.A. Altmann, S.S. Strickland and O.T. Oftedal (1991). Traditional diet and food preferences of Australian Aboriginal hunter-gatherers. Philosophical Transactions: Biological Sciences 334(1270): 233–41.
Oliveira, N.V. (2008). Subsistence archaeobotany: food production and the agricultural transition in East Timor. Doctoral thesis, Australian National University, Canberra, ACT.
Ouzman, S., P. Veth, C. Myers, P. Heaney and K.F. Kenneally (2019). Plants before animals?: Aboriginal rock art as evidence of ecoscaping in Australia’s Kimberley. In B. David and I.J. McNiven, eds. The Oxford handbook of the archaeology and anthropology of rock art, 1–13. Oxford: Oxford University Press.
Owen, T., J. Field, S. Luu, P. Kokatha Aboriginal, B. Stephenson and A.C.F. Coster (2019). Ancient starch analysis of grinding stones from Kokatha Country, South Australia. Journal of Archaeological Science: Reports 23: 178–88.
Pascoe, B. (2014). Dark emu: black seeds: agriculture or accident? Broome: Magabala Books.
Pawlik, A.F., P.J. Piper, M.G.P.G. Faylona, S.B. Padilla, J. Carlos, A.S.B. Mijares et al. (2014). Adaptation and foraging from the terminal Pleistocene to the early Holocene: excavation at Bubog on Ilin Island, Philippines. Journal of Field Archaeology 39(3): 230–47.
Pearsall, D.M. (2010). Paleoethnobotany: a handbook of procedures. California: Left Coast Press.
Perera, N., N. Kourampas, I.A. Simpson, S.U. Deraniyagala, D. Bulbeck,
J. Kamminga et al. (2011). People of the ancient rainforest: Late Pleistocene foragers at the Batadomba-lena rockshelter, Sri Lanka. Journal of Human Evolution 61(3): 254–69.
Piperno, D.R. (2018). A model of agricultural origins. Nature Human Behaviour 2(7): 446–7.
Porr, M. and E. Vivian-Williams (2021). The tragedy of Bruce Pascoe’s Dark Emu. Australian Archaeology 87(3): 300–4.
Popper, V.S. (1988). Selecting quantitative measurements in paleoethnobotany. In C.A. Hastorf and V.S. Popper, eds. Current paleoethnobotany: analytical methods and cultural interpretations of archeological plant remains, 53–71. Prehistoric Archeology and Ecology Series. Chicago: University of Chicago Press.
Reed, C.A., ed (1977). Origins of agriculture. The Hague: Mouton Publishers.
Roberts, P., A. Buhrich, V. Caetano-Andrade, R. Cosgrove, A. Fairbairn, S.A. Florin et al. (2021). Reimagining the relationship between Gondwanan forests and Aboriginal land management in Australia’s “wet tropics” iScience 24(3): 102190.
Rosenfeld, A. (1997). Excavation at Buang Merabak, central New Ireland. Bulletin of the Indo-Pacific Prehistory Association 16: 213–24.
Russell-Smith, J. (1985). Studies in the jungle: people, fire, and monsoon forest. In R. Jones, ed. Archaeological research in Kakadu National Park, 241–67. Canberra: Australian National University.
Russell-Smith, J., D. Lucas, M. Gapindi, B. Gunbunuka, N. Kapirigi, G. Namingum, K. Lucas, P. Giuliani and G. Chaloupka (1997). Aboriginal resource utilization and fire management practice in western Arnhem Land, monsoonal northern Australia: notes for prehistory, lessons for the future. Human Ecology 25(2): 159–95.
Sayok, A.K. and U. Teucher (2018). Loss of food plants knowledge and identity among Indigenous peoples in Malaysia. Journal of Advanced Research in Social and Behavioural Sciences 11(1): 174–88.
Scarlett, N.H. (1985). A preliminary account of the ethnobotany of the Kija people of Bungle Bungle outcamp. East Kimberley Working Paper 6. Canberra: Center for Resource and Environmental Studies.
Sievers, C. (2006). Seeds from the Middle Stone Age layers at Sibudu Cave. Southern African Humanities 18(1): 203–22.
Simchoni, O. and M.E. Kislev (2011). Early finds of Celtis australis in the southern Levant. Vegetation History and Archaeobotany 20(4): 267–71.
Smith, B.D. (2001). Low-level food production. Journal of Archaeological Research 9(1): 1–43.
Smith, M. (1982). Late Pleistocene Zamia exploitation in southern Western Australia. Archaeology in Oceania 17(3): 117–21.
Smith, M. and A.C. Kalotas (1985). Bardi plants: an annotated list of plants and their use by the Bardi Aborigines of Dampierland, northwestern Australia. Records of the Western Australian Museum 12(3): 317–59.
Spengler III, R.N. (2020). Anthropogenic seed dispersal: rethinking the origins of plant domestication. Trends in Plant Science 25(4): 340–48.
Sutton, P. and K. Walshe (2021). Farmers or hunter-gatherers? The Dark Emu debate. Melbourne: Melbourne University Press.
Ucko, P.J. and G.W. Dimbleby, eds (1969). The domestication and exploitation of plants and animals. London: Gerald Duckworth & Co. Ltd.
Vigilante, T., S. Ondei, C. Goonack, D. Williams, P. Young and D.M.J.S. Bowman (2017). Collaborative research on the ecology and management of the “Wulo” monsoon rainforest in Wunambal Gaambera Country, north Kimberley, Australia. Land 6(4): 68–88.
Wallis, L.A. and S. O’Connor (1998). Residues on a sample of stone points from the west Kimberley. In R. Fullager, ed. A closer look: recent Australian studies of stone tools, 149–78. Sydney: Archaeological Computing Laboratory, School of Archaeology, the University of Sydney.
Walsh, M. (2021). There’s something about marnunggurrun: wood charcoal as an indicator of earth ovens and anthropogenic fire regimes at Walanjiwurru 1, Marra Country. Honours thesis, University of Western Australia, Crawley, WA.
Walsh, M., E. Dotte-Sarout, L.M. Brady, J. Bradley, J. Ash, D. Wesley, S. Evans and D. Barrett (2024). Collaborative anthracology and cultural understandings of wood charcoal in Marra Country (northern Australia). Archaeological and Anthropological Sciences 16(9): 148. DOI: 10.1007/s12520-024-02052-y.
Watson, P.J. and M. Kennedy (1991). The development of horticulture in the eastern woodlands of North America: women’s role. In J. Gero and M. Conkey, eds. Engendering archaeology, 255–75. Oxford: Basil Blackwell.
Wedage, O., P. Roberts, P. Faulkner, A. Crowther, K. Douka, A. Picin et al. (2020). Late Pleistocene to early-Holocene rainforest foraging in Sri Lanka: Multidisciplinary analysis at Kitulgala Beli-lena. Quaternary Science Reviews 231: 106200. DOI: 10.1016/j.quascirev.2020.106200.
Whitau, R. (2018). Late Quaternary human-environment interaction in Bunuba and Gooniyandi country, Western Australia. Doctoral thesis, Australian National University, Canberra, ACT.
Whitau, R., J. Balme, S. O’Connor and R. Wood (2017). Wood charcoal analysis at Riwi cave, Gooniyandi country, Western Australia. Quaternary International 457: 140–54.
Whitau, R., I.E. Dilkes-Hall, D. Vannieuwenhuyse, S. O’Connor and J. Balme (2018a). The curious case of Proteaceae: macrobotanical investigations at Mount Behn rockshelter, Bunuba Country, Western Australia. Australian Archaeology 84(1): 19–36.
Whitau, R., D. Vannieuwenhuyse, E. Dotte-Sarout, J. Balme and S. O’Connor (2018b). Home is where the hearth is: anthracological and microstratigraphic analyses of Pleistocene and Holocene combustion features, Riwi Cave (Kimberley, Western Australia). Journal of Archaeological Method and Theory 25(3): 739–76.
Yen, D.E. (1988). Introduction to group of papers about plants. In B. Meehan and R. Jones, eds. Archaeology with ethnography: an Australian perspective. Canberra: Australian National University.