5
Ulna
The ulna is generally the smaller of two paired bones that make up the fore limb (the other being the radius). It is responsible for flexion and extension, and forms a hinge with the humerus. In many species, the ulna is lateral to the radius, however in ungulates (hoofed animals), where it is often fused to the radius, it is posterior.
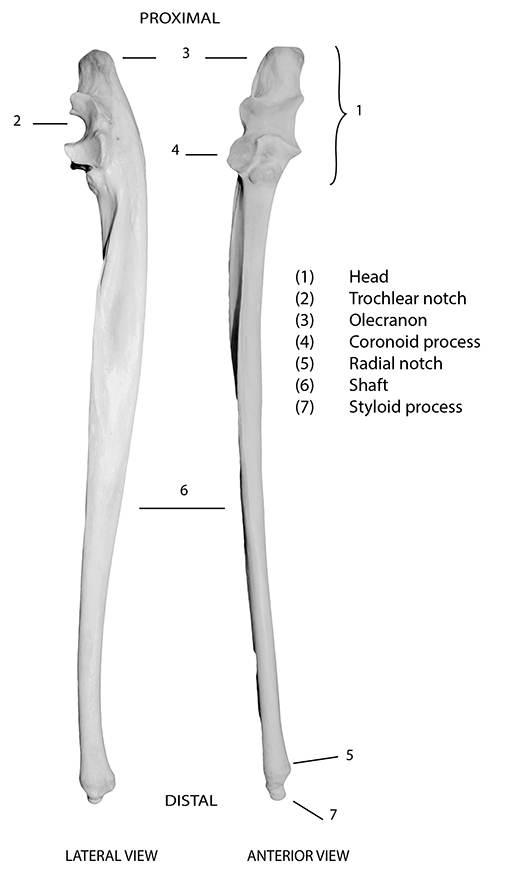
Figure 5.1: Kangaroo ulna with diagnostic features labelled.
Diagnostic features
Diagnostic features of the ulna are shown in Figure 5.1. When complete, the ulna is easily recognised by its hook-like head (1), which is comprised of a trochlear (semilunar) notch (2) for articulation with the trochlea of the distal humerus, and often ‘swollen-looking’ proximal cap, the olecranon process (3). The coronoid process (4) projects from the distal portion of the trochlear notch, and fits into the coronoid fossa of the humerus. Some species also have a radial notch (5), projecting slightly on the lateral side of the trochlear notch, for articulation with the radius. The ulna has a thin, tapering shaft (6) that ends in a pointed styloid process (7) in many species. In adulthood, the ulna is fused to the radius in most ungulates, especially horses, cattle, sheep and often pigs (Figure 5.2). In these species, fusion results in a greatly reduced ulnar shaft. As for the radius, many of the images on the following pages depict both elements in these species. Note that in species with a fused radius/ulna, the two bones are generally separate in immature animals. Since the radius enables rotation of the forearm, in species in which it is fused to the ulna, no rotation of the fore limb is possible.
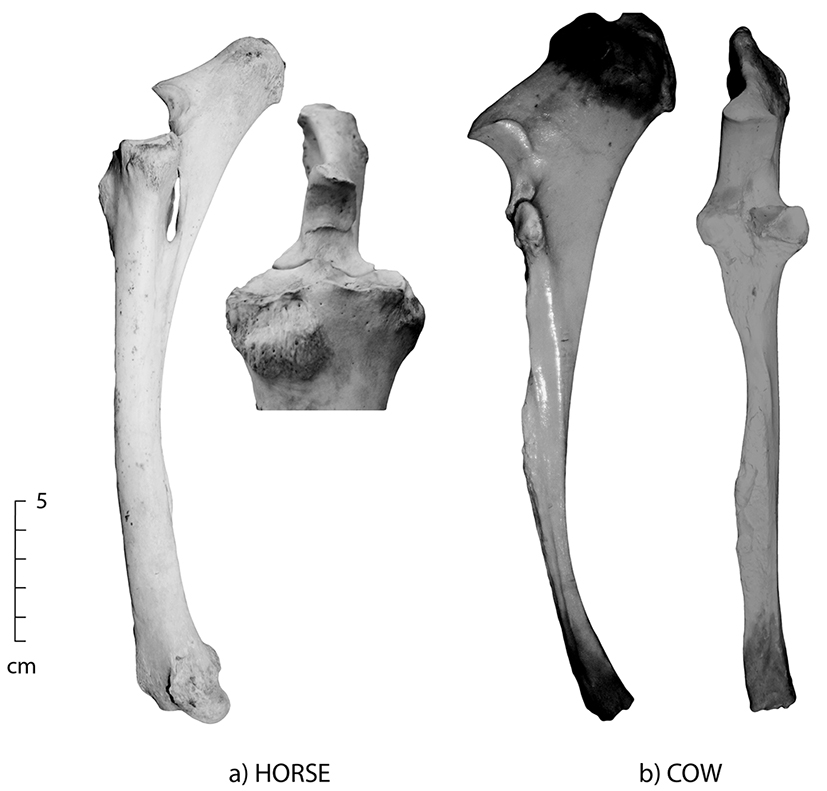
Figure 5.2: Lateral and anterior views of fused ulna/radius; (a) horse and (b) cow. The ulna and radius are fused in most adult ungulates.
Orientation and siding
To orient the ulna, the trochlear notch and olecranon process will be proximal and anterior (pointing up and forward), and the thin shaft will taper distally. In species with a distinct olecranon process, the curve can be used to indicate from which side the bone came. To side the ulna in this way, hold it with the trochlear notch facing you, and the olecranon process will ‘curve’ to the side from which it came. If the olecranon process is missing or less well developed, the morphology of the trochlear notch can be used. In this case (still holding the ulna with the trochlear notch facing you), the coronoid process will flare out on the side from which it came. Alternatively, you can hold the ulna in correct anatomical position, with the trochlear notch facing anteriorly, and side it by ensuring that the radial notch (if present) is lateral.
Species identification
Refer to Figures 5.2–5.6 for species identification using the ulna.
Distinguishing between human and macropod
The trochlear notches of both humans and macropods appear quite similar and the ulna of both species is rather long and thin. To differentiate between the two, it is often easiest to examine the proximal and distal ends of the ulna.
- Humans do not have a distinctive, well-developed olecranon process, but do have a slightly flared distal end right above a distinctive styloid process.
- Macropods have a well-developed olecranon process and a tapered (as opposed to flaring) distal end.
- Humans have a distinct sharp crest along the length of the ulnar shaft, which is only extant on the mid-shaft of macropods.
- In humans, the ulna has a teardrop-shaped mid-shaft section and is long and thin (Figure 5.3a).
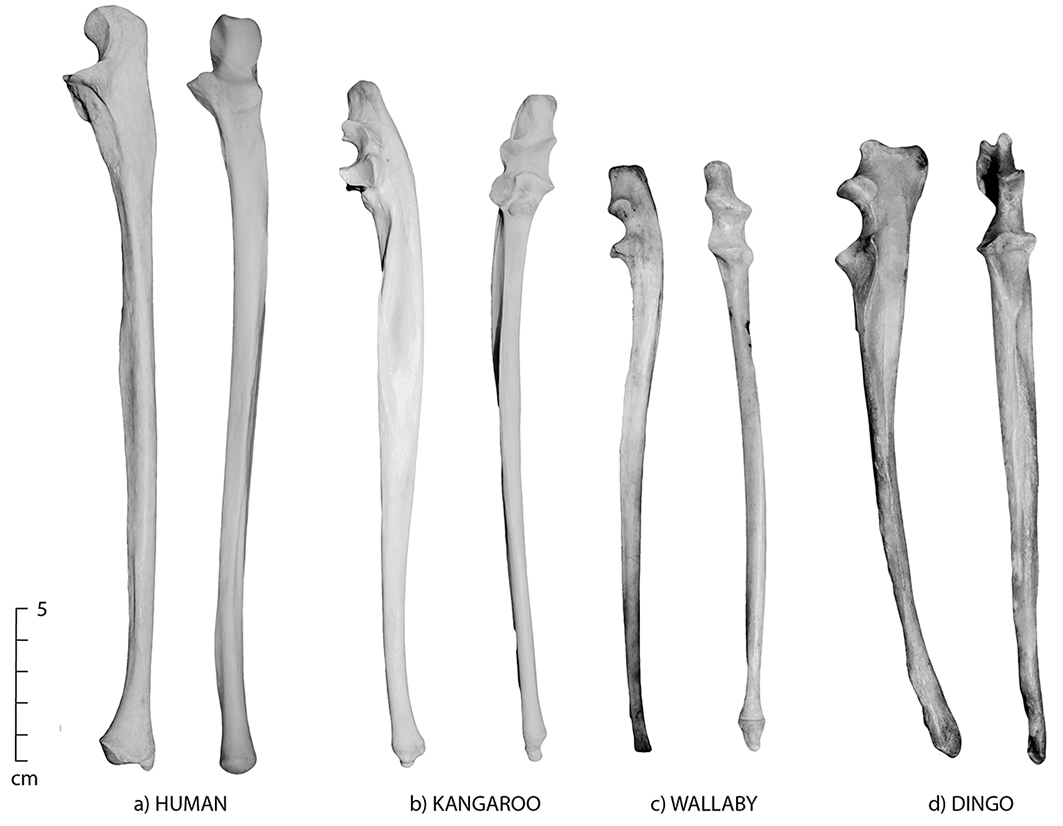
Figure 5.3: Lateral and anterior views of the ulna; (a) human, (b) kangaroo, (c) wallaby and (d) dingo. Note the close similarities but distinct size differences of the four species. The human ulna has a teardrop cross-section and is long and thin.
Differences between non-human species
- The ulna is distinguished by a ‘flattened’ appearance medio-laterally, with a shaft that becomes narrower distally.
- The ulna and radius are fused in most adult ungulates (horses, cattle, sheep and pigs).
- The distal ulna is severely reduced in most ungulates, especially horse.
- In species in which the radius and ulna do not fuse (humans, dingoes, cats, macropods, possums, quolls, bandicoots, wombats and rabbits), the ulnar shaft may be confused with a medial fibula shaft fragment.
- In adult horses, the ulna is so reduced in length the distal end tapers to a point that does not extend the full body of the radius.
- In dingoes and cats, the ulna has a prominent bulging mid-shaft tuberosity.
- To distinguish between cats and small dingoes, the distal ulna is rather small in dingoes and more prominent in cats; dingoes have a high olecranon process (distally) (Figure 5.4).
- Horses, cattle, sheep, pigs, macropods and wombats all have long and massive curved olecranon processes (Figure 5.5).
- Kangaroos, wallabies and possums all have very similar ulnar morphology. Use the size and curvature of the shaft to make a general distinction between most species of kangaroo and wallaby (kangaroos have a more pronounced anterior curve than wallabies) (Figure 5.3b and c).
- To differentiate between possums and wallabies, size may be used (wallabies are slightly larger), as well as robustness of the ulnar shaft. In general, the shaft is slightly more robust as it nears the distal end in possums, and more tapered in wallabies (Figures 5.6b and 5.3c).
- To distinguish between rabbits and possums, look at the shaft – rabbit ulnae curve anteriorly and have a deeper trochlear notch, whereas possums have a relatively straight shaft and a shallow trochlear notch (Figures 5.6a and b).
- To distinguish between wombats and pigs, look at the coronoid process, which flares outward in wombats (Figures 5.5b and c).
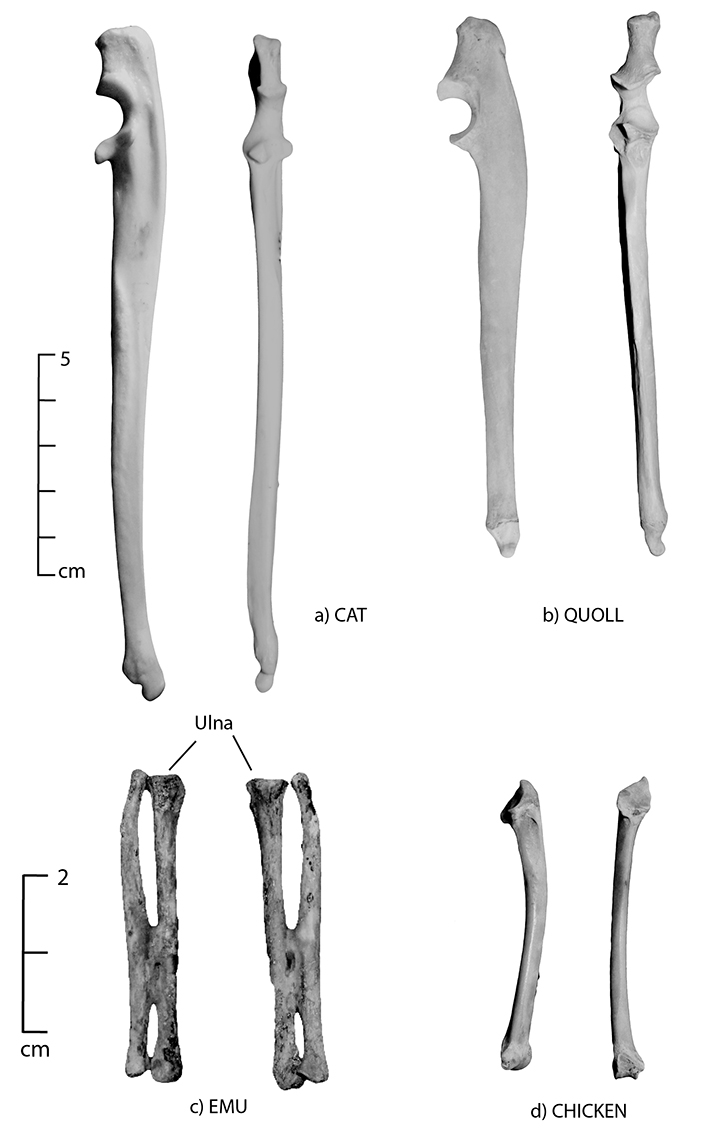
Figure 5.4: Lateral and anterior views of the ulna; (a) cat, (b) quoll, (c) emu and (d) chicken. The dingo (Fig. 5.3d) and cat ulna have a prominent and bulging mid-shaft tuberosity. To differentiate between the two, the distal portion of the dingo ulna is small, while that of the cat has a more prominent distal end.
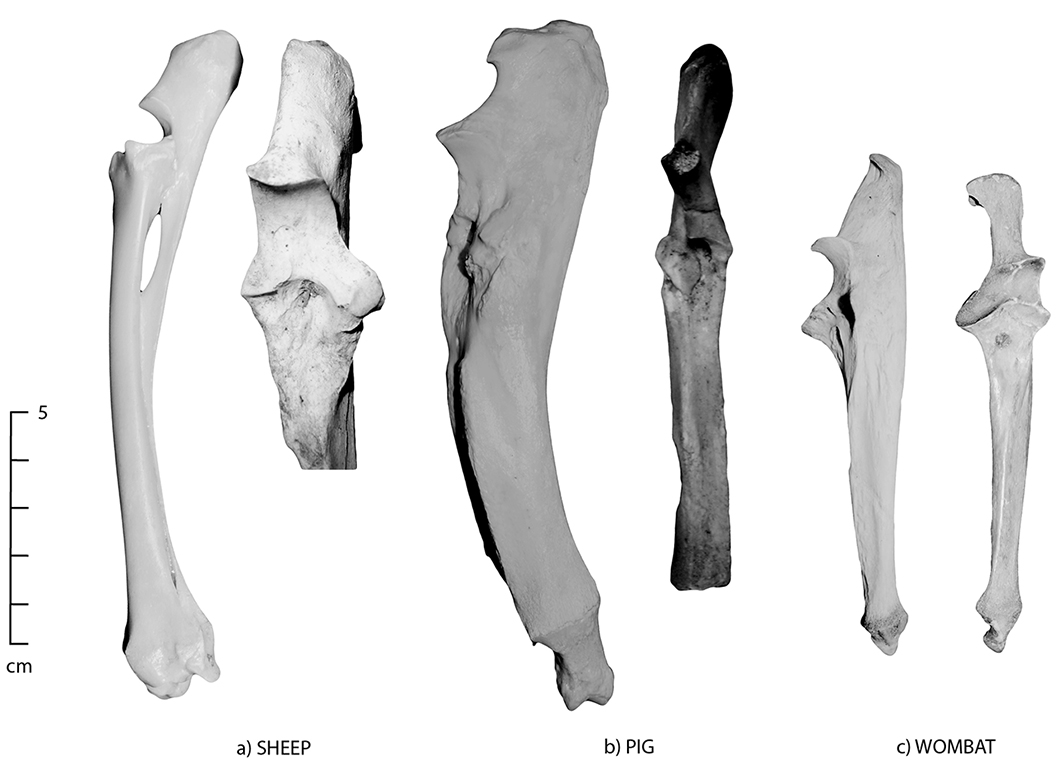
Figure 5.5: Lateral and anterior views of the ulna; (a) sheep, (b) pig and (c) wombat. The pig has a very large and curved olecranon, which is similar in morphology to wombat.
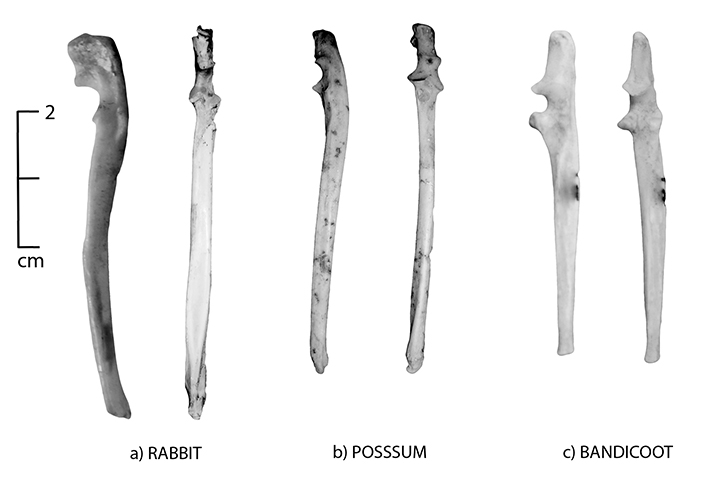
Figure 5.6: Lateral and anterior views of the ulna; (a) rabbit, (b) brushtail possum and (c) bandicoot.
Common state in archaeological assemblages
The ulna is frequently encountered in archaeological assemblages; however, it is very rarely complete. Generally, the relatively robust trochlear notch is extant, and can be used to identify species. The shaft is often broken, and even if extant, it is likely to be unidentified as it may be mistaken for a rib or fibula fragment. In species with a fused radius/ulna, part of a broken ulnar shaft may be extant on the posterior radial surface. The olecranon process is also often missing, as its proximal end is a fusion centre, and has an epiphyseal cap that may not be extant. This is especially prevalent in those species in which the olecranon process is more exaggerated and swollen-looking (i.e. horses, cattle, sheep, pigs, kangaroos and wallaby). In pigs, the ulna is generally found unfused in most historical archaeological assemblages due to the relatively young age at which pigs are slaughtered.
In species utilised for meat, primary butchery of a carcass tends to make an initial cut just above, or at, the articulation of the radius/ulna and humerus to disarticulate the upper from the lower fore leg. This often results in a cut mark through the trochlear notch. As is the case with the radius, the ulna is often left articulated with the humerus to form a leg cut (e.g. ham or lamb) that may be cooked with the bone. In rabbit, the ulna is often intact as rabbits are frequently not butchered prior to cooking; the same pattern is prevalent in poultry.
The reduced ulnae of some species are especially prone to different preservation and other taphonomic factors, such as scavenging. For example, emu ulnae are rarely found intact, as the wing is one of the first body parts to be scavenged. In macropods such as kangaroos and wallabies, there is little meat on this element, and it is commonly left behind by people interested in higher meat-bearing elements. In naturally accumulated assemblages, macropod ulnae are commonly found articulated with the entire hand or wrist (ulna, radius, carpals, metacarpals and phalanges), as even scavengers tend to ignore this element. The same pattern is commonly encountered in many species.
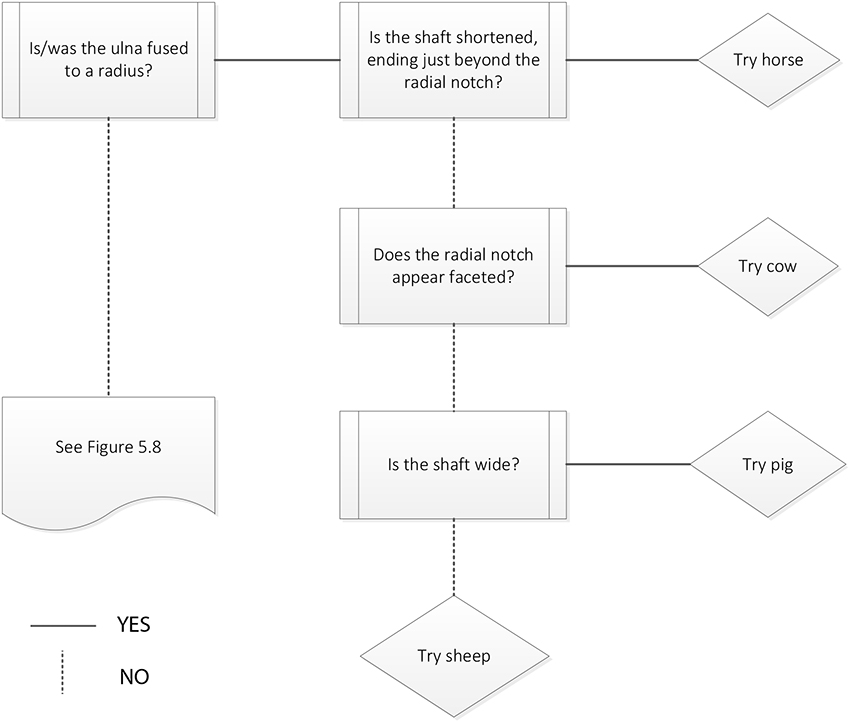
Figure 5.7: Ulna decision process 1. Ulna fused to radius.
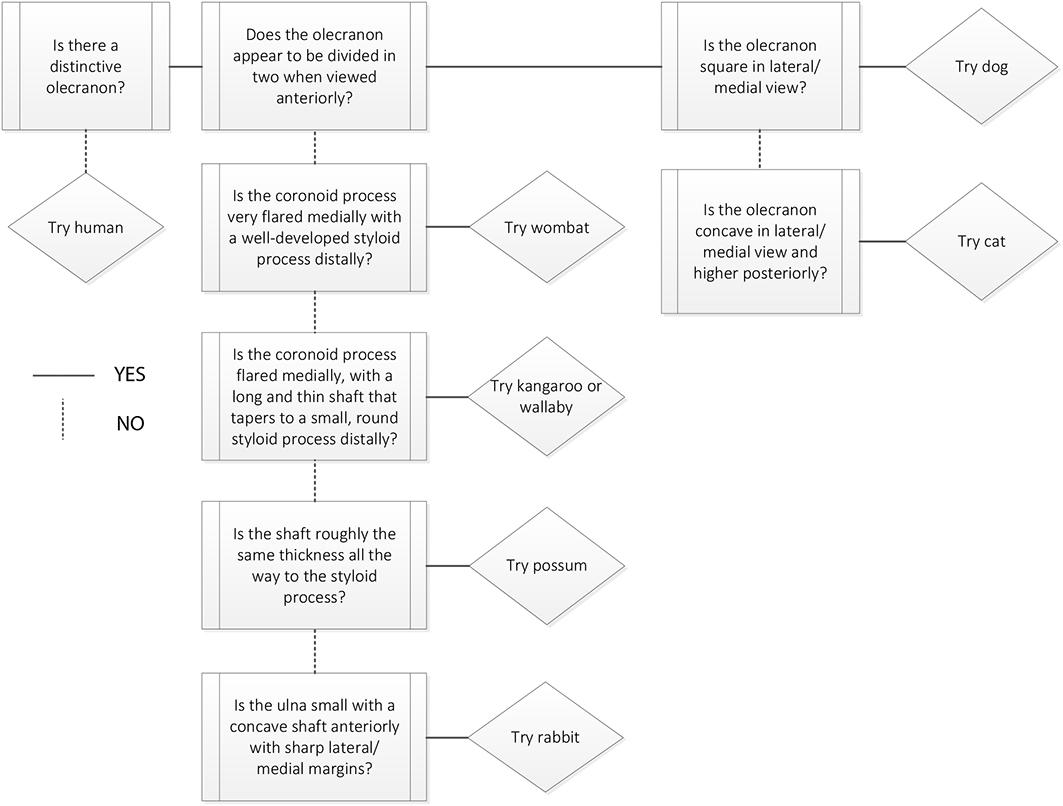 Figure 5.8: Ulna decision process 2. Ulna and radius not fused. |